Теории цветового зрения - концепции, объясняющие способность чело…») при совместной работе с Александровым над матермалом Теории цветового зрения с последующей моей работотой в Теории цветного зрения. Было принято решение для создания двух статей для сравнения материалов и в итоге для создания одной статьи.[2]. В Науке я работал параллельно. В настоящее время я корректирую и дополняю материал моей статьи Теории цветного зрения.
Подготовка материала для объединения и согласования текста двух близких статей:[3].
Теории цветового зрения — авторская статья Александрова. (11:34, 26 января 2010 Alexandr (обсуждение | вклад) . . (13 869 байт) (+13 869) . . (новый стаб - ответвление от Зрение человека)).
Теории цветного зрения — авторская статья Мига. (08:37, 10 апреля 2010 Миг (обсуждение | вклад) . . (1613 байт) (+1613) . . (← Новая: {{{Пишу}}} Теории цветного зрения — каждая теория цветного зрения может считаться действи…)).
В 22:04, 26 марта 2012 Миг (обсуждение | вклад) м . . (103 200 байт) (0) . . (переименовал Участник:Миг/Теории цветового зрения в Участник:Миг/Теории цветного зрения: Уточнение названия)[4].
Ещё 11:04, марта 27, 2012 137vai (обсуждение | вклад) м . . (103 107 байт) (+49 437) . . (Правки DmitriyRDS (обсуждение) откачены к версии Moisey). (См. историю). Статья создана мною впервые с Александровым. [5].
17:30, 29 апреля 2015 Serebr (обсуждение | вклад) м . . (59 763 байта) (0) . . (Serebr переименовал страницу Теории цветового зрения (версия DmitriyRDS) в Теории цветового зрения поверх перенаправления: утверждаю эту ве…). В Традиции сейчас записано Теории цветового зрения со сноской (Serebr): См. также Теории цветного зрения (версия Миг), Теории цветового зрения (версия DmitriyRDS).
Теории цветного зрения — концепции, объясняющие способность человека различать цвета, основанные на наблюдаемых фактах, предположениях, их экспериментальной проверке. Теории нередко имеют предсказательную силу, помогая предсказать дотоле не наблюдаемые эффекты.
В истории есть многие наивные теории, предсказательная сила которых была невелика. Даже гениальные предположения учёных прошлого были в основном умозрительны, так как естественные науки прошлого не обладали необходимым инструментарием; не был сформирован понятийный аппарат, невозможно было провести многие критически важные эксперименты. Бурное развитие естественных наук можно отнести к ХIХ веку, когда специалисты в области биологии, химии и физики продвинули далеко вперёд науку цветного зрения в области естественных наук.
В идеале теория обладает силой предвидения, позволяя обнаружить новые особенности явлений; именно новая теория полезна тогда, если она объясняет известные эффекты более старой теории, но более ясно, или с меньшим числом предположений. На сегодняшний день доказано, что цветное зрение основано и функционирует на базе трихроматизма. (Cм. Теория многокомпонентного цветного зрения).
Теории цветного зрения, Теория многокомпонентного цветного зрения или Теория трёхкомпонентного цветного зрения созданы на базе физики цвета (RGB) и взаимодействия с ним живой среды обитания — биологической, а также на основе колориметрических, гистологических, генетических исследований,[1] на базе достижений в этой области более, чем за 30‒35 лет, на основе биофизики, биохимии созданы с учётом пересмотра основ прежнего сложившегося процесса цветного зрения с разных точек зрения:
С точки зрения биологической, в области цветного зрения начиная с 1966 по 2009 годы (Труды доктора Р.Е.Марка и его лаборатории) с основными экспериментальными данными исследований живой клетки, на срезах сетчатки установлена работа колбочек и (S,M,L) в блоке RGB и палочек.[2] В условиях дневного освещения (цветного зрения) — колбочек и работа палочек, в период сумеречного и ночного освещения (не цветного) изолировано от колбочек. Работа фоторецепторов связана с полученными данными работы видоизменяющихся разновидностей фотопигментов на базе белков опсинов:
С точки зрения чисто физической на базе труда (2011 года) учёного физика Джеральда К. Хата[4], который на базе физики взаимодействия света с внешними долями мембран фоторецепторов сетчатки глаза, где в основном осуществляется первичное взаимодействие со светом на наноуровне («nanostructural») (рецепторное) и доктора физика Джона Медейроса,[5] который рассмотрел работу внешних долей мембраны колбочек и палочек с точки зрения аналогичной работе волноводов конической и цилиндрической формы в среде прозрачного тела глаза (жидкая среда). В конечном итоге, физики Дж. К. Хат и Джон Медейрос пришли к общепринятому принципу трихроматизма. (См. Пересмотр традиционных взглядов на зрительный процесс физика К. Хата, Работа внешних мембран колбочек и палочек сетчатки глаза как волновод).
Данные доктора, физика Джона Медейроса, рассмотрев внешние доли мембран колбочек и палочек, он их рассматривает как биологические волноводы, которые воспринимают световые лучи с фиксацией их в сечениях волноводов, равных размерам поперечного сечения фронта световой волны в обратном порядке прохождения их (внешних долей мембран колбочек) в жидкой среде, нежели в обычных оптических волноводах в воздухе. (Связано с влиянием показателей преломления сред).
Фундаментальные исследования академика РАН биохимика М. А. Островского[6] в области работы фотопигмента родопсинаБиохимия зрения и свободно-радикальное окисление клеток сетчатки глаза на базе которых было также подтверждено, что палочки с фотопигментом родопсином участвуют в зрительном процессе в условиях сумеречного и ночного зрения (чёрно-белого);
То, что мы ощущаем как цвет, представляет собой комбинированное воздействие:
Спектрального распределения светового потока из дающего энергию источника света;
Физических и/или химических свойств всех материалов, пропускающих или отражающих световой поток (по меньшей мере часть светового потока, переориентированную в сторону глаза);
Физиологической реакции глаза на световой поток, включающей в себя нервные импульсы, передаваемые в зрительные отделы головного мозга;
Влияние специфики работы мембраны колбочек при цветном зрении.
Переработки нашим мозгом этих сигналов в сочетании с сигналами из соседних областей поля зрения, нашими воспоминаниями о сходных ситуациях, имевших место в прошлом опыте.[7] Хотя это достаточно общее высказывание в целом верно, но для того, чтобы «теория цвета» стала продуктивной, требуется дальше развить и углубить, связать с фактами каждую модель уровней цветовосприятия.
Общие требования[]
Теория цветного зрения как и любая теория может считаться эффективной только при выполнении следующих условий:
теория должна строиться на объективных экспериментальных данных:
данных с использованием результатов микроскопии и гистологического анализа (например, люминесцентной микроскопии живых тканей),
учёт работы фоторецепторов колбочек, с точки зрения влияния конической формы внешней доли мембраны, работы её как волновода конической формы в жидкой среде, наличия классов колбочек, меняющие свои размеры, и характеристики, связанные с градиентом уменьшения диаметра большего сечения конуса с одновременным увеличением длины мембраны с центробежным вектором от центра жёлтого пятна к периферии сетчатки. (Феномен, открытый учёным Джон А. Медеиросом, см. Специфика работы мембраны колбочек),
учёт различных симптомов всевозможных видов цветовой слепоты,
основываться на биофизических и биохимические законах,
описываться математическими зависимостями в трёхмерном 3D пространстве.[8]
Исходить из первых шагов исследований учёных Юнга-Гельмгольца и Эвальда Геринга двух основных теорий цветного зрения.
Две основные теории цветного зрения[]
Трихроматическая теория, также известная как теория Юнга-Гельмгольца;
Теория процесса оппонентного отбора основных цветов Эвальда Геринга RGB.
Эти две теории дополняют друг друга и объясняют процессы, которые работают на разных уровнях визуальной системы.
Трихроматическая теория[]
Доказательства для трихроматической теории вытекают из сопоставления цветов и исследований смешивания цветов (например, как художник смешивает краски). Янг и Гельмгольц провели эксперименты, в ходе которых индивидуумы отрегулировали относительную интенсивность 3 источников света различной длины волны, так, что полученное поле смеси соответствовало соседнему испытательному полю, состоящему из одной длины волны. Для индивидуумов с нормальным цветовым зрением необходимы три разных длины волны (т. Е. Праймериз) для соответствия любой другой длине волны в видимом спектре. Это открытие привело к гипотезе о том, что нормальное цветовое зрение основано на активности трех типов рецепторов, каждый из которых имеет различную пиковую чувствительность. В соответствии с трихроматической теорией мы теперь знаем, что общий баланс активности в S (короткая длина волны), M (средняя длина волны),
S явление, (everal) современная тема восприятия цвета не может быть объяснена только в трехцветной теории. Однако, например, индивидум не может учитывать дополнительные наборы цветовых образцов, в которых расширенный осмотр одного цвета приведет к последующему восприятию его дополнительного цвета. (См. ниже). Дополнительные явления слепоты лучше объясняются теорией процесса оппонентного отбора цветов..
Теория процесса оппонентного отбора цветов[]
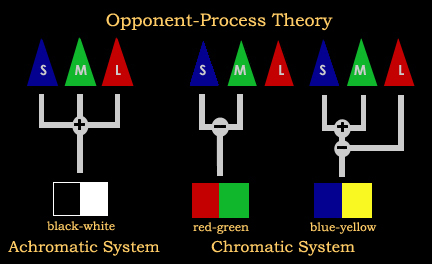
Разработанная Эвальдом Герингом (1920/1964), теория оппонентного процесса утверждает, что конические фоторецепторы связаны между собой, образуя три противоположные цветовые пары: синий / желтый, красный / зеленый и черный / белый. Активация одного члена пары ингибирует, подавляет активность в другом. В соответствии с этой теорией, два члена пары не могут быть замечены в одном месте, что объясняет, почему мы не испытываем таких цветов, как «голубовато-желтый» или «красновато-зеленый». Эта теория также помогает объяснить некоторые виды дефицита цветового зрения. Например, люди с дихроматическими недостатками могут соответствовать тестовому полю, используя только два первичных элемента. В зависимости от недостатка они будут путать красный, зеленый или синий и желтый.
Теория оппоненного процесса объясняет, как мы видим желтый, хотя желтой колбочки нет. Это связано с возбуждающими и тормозящими связями между тремя типами колбочек. В частности, суммируется одновременная стимуляция красного (L колбочек) и зеленого (M колбочек) и, в свою очередь, блокирует B + Y- (синий-жёлтый), что приводит к восприятию желтого. Однако, когда присутствует синий свет, активируется S-кодбочка, ячейка B + Y- принимает возбуждающий вход, а синий воспринимается.
Теория оппонентного процесса.
Можно видеть отношения противника между красным и зеленым, а также синим и желтым. Просмотрите четырехцветные исправления послеожогового пятна ниже в течение 30 секунд. Затем удалите цветовые стимулы, наведя курсор мыши на изображение, заставив его стать пустым белым полем. Когда идет фиксация в точке в центре поля, вы должны заметить, что исходные цвета меняются на противоположные — там, где вы видели красный цвет, теперь зеленый и наоборот. Аналогично для синего и желтого.
Цветовые оттенки с переходом а белый цвет (см. текст).
Дополнительный следящий. (Смотри белый цвет в атаре [6])
Как цветовые оттенки объясняются теорией оппонентного процесса? Когда один член цветовой пары «устал» посредством расширенного контроля, уменьшается ингибирование его соответствующего парного члена. Это увеличивает относительный уровень активности члена неустойчивой пары и приводит к восприятию цвета.
Трихроматическая теория или теория оппонентного процесса?
Фактически, как вы видели, обе теории необходимы, чтобы объяснить, что известно о цветовом видении. Трихроматическая теория объясняет явления цветного зрения на уровне фоторецепторов, а теория оппонентного процесса объясняет феномены цветного зрения, которые возникают из-за того, что фоторецепторы взаимосвязаны при помощи нервной системы.
История[]
Религиозная гипотеза[]
Человека всегда волновали вопросы связанные с работой органа зрения. И во многих религиях даётся своё описание устройства глаза и принципа зрения человека. Всё как правило сводится к тому, что глаз человека является настолько сложным и совршенным органом, что его не могла создать природа в процессе эволюции, откуда, система зрения четко свидетельствует о существовании Бога-Творца.
Воззрения древнего мира[]
Древнегреческие философы 6-4 вв. до н.э.: Эмпедокл, Демокрит, Аристотель о цветном зрении.Первыми исследователями, положившими начало науки «о цвете», были древнегреческие философы 6-4 вв. до н.э.: Эмпедокл, Демокрит, Аристотель. Непосредственные наблюдения и размышления о явлениях и ощущении цвета позволили сделать чисто эмпирические выводы о цвете. Цвет был разделен по категориям, искалось решение проблемы о происхождении цвета и цветовых явлений.
Аристотель попытался упорядочить все многообразие цвета и цветовых тонов, расположив видимые цвета между белым и черным цветом, от положительно воспринимаемых до отрицательно воспринимаемых, как по вкусовым характеристикам от сладкого до кислого. Уже тогда цвета были
разделены на основные и производные от других. Аристотель и Демокрит высказывали гипотезу о существовании глазных лучей, способствующих
восприятию цвета, и считали, что свет исходит из самих тел, которые нами воспринимаются.
Вопрос: «Как появляются цвета и какие цвета являются производными» волновал художников, философов и ученых долгое время. Постоянно шли споры о том, какая теория цвета – «трех» или «четырехкомпонентная», является верной и правильной.
Сторонниками «четырехкомпонентной» теории образования цвета были: Эмпедокл (530 – 430 гг. до н.э.), Леонардо да Винчи (1452 – 1519), И.В. Гете (1749 – 1832) и другие. По данной теории, существуют 4 основных цвета, образующие другие цвета в соответствии с четырьмя основными элементами Вселенной, которые выделяются на основании феноменологических данных.
В «трехкомпонентной» теории главными являются 3 основных цвета. «Трехкомпонентной» теории образования цвета придерживались такие ученые – исследователи, как: Тобиас Майер (1723 – 1762), Иоганн Генрих Ламберт (1728 – 1777), Ф.О. Рунге (1777 – 1810), В. Освальд (1853 – 1932) и другие. Этим спорам не положило конец даже открытие «волновой теории цвета». Прародителем «волновой теории цвета» стал итальянский физик и астроном Ф.И. Гримальди (1618 – 1663), открывший эффект призматического спектра. Дифракция света (дифракцией света – преломление луча света на кристалле и разложение его на цветовые лучи) была описана им в труде «Физико-математический трактат о свете, цветах и радуге».[9]
То есть, еще философы древнего мира говорили об основных цветах и о трихроматизие!
Полтора столетия спустя, в 1672 году, увидел свет первый капитальный труд по теории цвета. Назывался он «Новая теория света и цвета» Ньютона. Пропустив солнечный свет через призму, Ньютон разложил его в спектр, получив радугу. Он первым показал, что белый цвет всегда сложен. Основной вывод ученого стал фундаментальным для науки о цвете: « … и цвета относятся к физике, и науку о них следует почитать математической, поскольку она излагается математическим рассуждением». Однако он не учитывал биофизического механизма восприятия цвета и исходил из предположения, что цвет является свойством света. В качестве основных цветов Ньютон впервые предложил наиболее различающиеся цвета радуги: красный, оранжевый, жёлтый, зелёный, голубой, синий, фиолетовый. Поиски гармонии (аналогично тонам звуковой гаммы) привели и в теории цвета к появлению числа «семь».
Представление о биофизическом восприятии цвета в середине XVIII столетия впервые ввел М. В. Ломоносов (Л.). Это было его «Слово о происхождении света, новую теорию о цветах представляющее, июля 1-го дня 1756 года оговоренное», речь, произнесенная Л. на русском языке в публичном собрании Академии наук 1 июля 1756 г., в которой он изложил итоги своих долголетних теоретических и экспериментальных изысканий, связанных с природой света. Впервые С. опубликовано на русском языке отдельной брошюрой в Санкт-Петербурге в 1757 г. В 1759 г. по требованию Л. было издано на латинском языке в переводе Г. В. Козицкого. Рукопись работы, написанной в мае — июне 1756 г., не сохранилась. :[10]
"Три рода эфирных частиц имеют совмещение: «первой величины эфир с соляною, второй величины с ртутною, третьей величины с серною или горючею первоначальною материею». «Наконец, нахожу, что от первого рода эфира происходит цвет красный, от второго — желтый, от третьего — голубой. Прочие цветы рождаются от смешения первых».
Основные положения гипотезы Ломоносова:
количество основных цветов сведено к трем (красный, жёлтый, голубой) — это то минимальное число цветов, которые совмещаются (смешиваются) в различной комбинации, позволяя получить все цветовые тона (но это не первичные основные цвета RGB);
воздействие на глаз различно по характеру, но едино по своей природе («коловратное движение эфира» — где все частицы эфира в виде зубчатых колёс приводятся во вращение в силу зубчатого зацепления друг с другом);
оговорено участие в цветовом зрении трёх зон спектра.
Так в теориях цветового зрения появилось число «три» (например, три основных цвета, три колбочки, трихроматизм и др.). Гипотеза Ломоносова была первой, которая содержала основные требования, предъявляемые к теории.
Долгий спор Ньютона и Гёте[]
Теория цвета Иоганна Вольфганга Гёте, опубликованная в 1810 г., очаровывала физиков более ста лет. Гёте был не только поэтом, но и естествоиспытателем-энциклопедистом, с очень широким кругозором.
Гипотеза Т. Юнга[]
В 1802 году, спустя пол столетия после Ломоносова, Томас Юнг предположил, что глаз анализирует каждый цвет в отдельности и передаёт сигналы о нём в мозг по трём различным типам нервных волокон: один тип передаёт сигнал о наличии красного цвета, второй — зелёного, а третий — фиолетового.
Гипотеза Гельмгольца[]
Ещё пол-столетия спустя гипотезу Т. Юнга развил учёный Г. Гельмгольц, немецкий биолог и физик, который, впрочем, не упоминает известной работы Ломоносова «О происхождении света», хотя она была опубликована и кратко изложена на немецком языке.
Проведя серию опытов Гельмгольц сделал вывод, что для получения цветов требуется 4 или более основных цветов и, естественно полагал, что трёхкомпонентная гипотеза Юнга несостоятельна. Позже он предположил достаточность всего трёх основных механизмов исходя из предположения о том, что они обладают спектральной чувствительностью в широком, частично перекрывающемся диапазоне. Согласно предположениям его гипотезы в сетчаткеглаза человека должны быть три вида колбочек, максимум чувствительности которых приходится на красный, зелёный и синий участок спектра, то есть соответствуют трём «основным» цветам. Правда эта гипотеза не может объяснить ни механизм обработки сигналов, ни постоянство ощущения цвета (константность цвета) при изменении спектрального состава источника света.
В настоящее время сторонниками признанной теории многокомпонентного цветного зрения на базе экспериментов с применением современного флюоресцентного микроскопа. Они исследовали механизм цветного зрения на работе живых колбочек и палочек в сечениях сетчатки в цвете, в трёхмерном измерении. На мониторе микроскопа, получены данные топографии ячеек колбочек (например, наличие в каждой ячейке по четыре колбочки: где соответственно работают четыре колбочки и каждая из которых оппонентно выделяет независимо основные монолучи спектра:
фиолетовые,
синие,
зелёные,
красные.
Все эти данные получены при гистологическом исследовании колбочек, палочек у птиц. («четырехроматик»)[11],[12](См.Цветное зрение у птиц, 2009 год). (У человека и некоторых млекопитащих животных принята система «трихроматик» — восприятие цветов RGB).
Объединённая теория Юнга-Геринга[]
Объединённая теория Юнга-Геринга (Трихроматическая теория, или трицептивная теория) имела достаточно широкое распространение. Теории объясняет законы смешивания цветов, но не объясняет тот факт, что дихроматы не могут воспринять цвет фоторецептора, который отсутствует или деградирован. Например, у дальтоников колбочка-M (зелёный цвет) не развита или нарушена функция управляющего фоторецептора ipRGC или функция формирования цветного оптического изображения на нейронном уровне).
Гипотеза Эвальда Геринга[]
Карл Эвальд Константин Геринг (1834—1918) — немецкий физиолог.
Немецкий физиолог Эвальд Геринг изучал медицину. Работы Геринга касаются главным образом психофизики; кроме того, он исследовал пространственное чувство глаза, причем явился защитником нативистической школы в противоположность Гельмгольцу. Оспаривал психофизический закон Фехнера и установил новую теорию цветов. (Нативизм — теория восприятия пространства, по которой протяженность является неотделимой принадлежностью, прирожденным свойством наших ощущений, сознается нами как протяженное.[7])
Он саму сетчатку глаза наделял способностью пространственного (стерео) видения, по его мнению, восприятие глубины в условиях диспарантности (Диспаратность (от лат. disparatus — разделённый) варианта написания — когда различие взаимного положения точек, отображаемых на сетчатках левого и правого глаза.) обусловлено тем, что нейроны внутренней части сетчатки и внешней откалиброваны на восприятие разной степени глубины («Beitrage zur Physiologie», Lpz., 1861). Развивал теорию цветового зрения (1875), в которой цветоразличение объяснялось процессами диссимиляции (раздробление, изменение, разрушающее сходство, подобие) и ассимиляции (ассимиляция — совокупность процессов синтеза в живом организме), происходящими в трех типах клеток сетчатки (колбочек), ответственных за восприятие трех качеств (белое-черное, красное-зеленое, жёлтое-синие). В 1870 он впервые сформулировал гипотезу оппонентного цветного зрения. Он опирался не только на существование пяти психологических ощущений, но также и на тот факт, что они действуют в противоположных парах, одновременно дополняя и исключая друг друга. Т.к. «разные» цвета при смешивании образуют промежуточные цвета, например, зелёный и синий, жёлтый и красный. Другие пары промежуточных цветов образовать не могут, зато дают новые цвета, например красный и зелёный. Хотя красный и зелёный цвета дают жёлтый цвет. (Фовея сетчатки глаза видна как жёлтая, в ней практически колбочки M,L). Э.Геринг пришел к выводу, что таких пар цветов три: красный и зелёный, жёлтый и синий, белый и чёрный. Использование четырёх цветов при синтезе цвета дает больше возможностей, чем использование трёх. Модель Геринга хорошо объяснила «отрицательные» последовательные образы. Модель Эвальда Геринга обрела не только сторонников, но и противников. Доводы последних сводились в основном к следующему. Во-первых: пять разных типов светоприёмников в глазу — многовато. К тому же, зачем жёлтый рецептор, если жёлтый цвет получается смешением сигналов «красного» и «зелёного»? Во-вторых, почему противоположные жёлтый и синий дают белый цвет, а противоположные красный и зелёный — жёлтый?
Таким образом, теория Э.Геринга, развитая Харвичем и Джеймсоном, известна как Теория оппонентного цветного зрения. В ней сохраняется три системы рецепторов: красно-зеленые, желто-голубые и черно-белые. Предполагается, что каждая система рецепторов функционирует, как противоположная, антогонистическая пара. Также как и в теории Юнга — Гельмгольца, считается, что каждый из фоторецепторов (или пар фоторецепторов) чувствителен к свету волн разной длины, но максимально чувствителен к волнам определенной длины.
[8]
ХХ-XXI век — современные теории цветового зрения[]
Если яркое зелёное кольцо окружает серый круг, то последний в результате одновременного цветового контраста приобретает красный цвет. Явления одновременного цветового контраста и последовательного цветового контраста послужили основой для теории процесса оппонентных цветов или процесса противникаШаблон:,[13] предложенной в XIX в. Эвальдом Герингом. Геринг предполагал, что имеются четыре основных цвета — красный, жёлтый, зелёный и синий — и что они попарно связаны с помощью двух антагонистических механизмов — зелёно-красного механизма и жёлто-синего механизма.
Постулировался также третий оппонентный механизм для ахроматически дополнительных цветов белого и черного. Из-за полярного характера восприятия этих цветов Э.Геринг назвал эти цветовые пары оппонентными цветами. Из его теории следует, что не может быть таких цветов, как зеленовато-красный и синевато — жёлтый .
Таким образом, теория оппонентных цветов постулирует наличие антагонистических цветоспецифических нейронных механизмов.
Например, если такой нейрон возбуждается под действием зеленого светового стимула, то красный стимул должен вызывать его торможение.
Предложенные Эвальдом Герингом оппонентные механизмы получили частичную поддержку после того, как научились регистрировать активность нервных клеток, непосредственно связанных с рецепторами. Так, у некоторых позвоночных, обладающих цветовым зрением, были обнаружены красно-зелёные и жёлто-синие горизонтальные клетки. У клеток красно-зелёного канала мембранный потенциал покоя изменяется и клетка гиперполяризуется, если на ее рецептивное поле падает свет спектра 400‒600 нм, и деполяризуется при подаче стимула с длиной волны больше 600 нм. Клетки жёлто-синего канала гиперполяризуются при действии света с длиной волны меньше 530 нм и деполяризуются в интервале 530‒620 нм.
На основании таких нейрофизиологических данных можно составить несложные нейронные сети, которые позволяют объяснить, как осуществить взаимную связь между тремя независимыми системами колбочек, чтобы вызвать цветоспецифическую реакцию нейронов на более высоких уровнях зрительной системы.
В своё время между сторонниками каждой из описанных теорий велись жаркие споры. Однако сейчас эти теории можно считать взаимно дополняющими интерпретациями цветового зрения. В зонной теории Крисса, предложенной 80 лет назад, была сделана попытка синтетического объединения этих двух конкурирующих теорий. Уже в то время он утверждал, что трёхкомпонентная теория пригодна для описания функционирования уровня рецепторов, а оппонентная теория — для описания нейронных систем более высокого уровня зрительной системы.[14]
В настоящее время они на базе последних открытий всё больше утверждаются. То есть, если считалось, что трёхкомпонентная теория цветного зрения рассматривает восприятие цвета только на уровне фоторецепторов сетчатки, а оппонентная теория служит для описания нейронных систем более высокого уровня зрительной системы (с участием головного мозга), то сейчас уже однозначно принято рассматривать цветное зрение на уровне общей трёхкомпонентной теории, неразрывно связанной с работой низших отделов зрительной систкмы — сетчаткой глаза (рецепторный уровень) во взаимодействии с головным мозгом (нейронный уровеь).[15]
Зонная теория Крисса[]
В свое время между сторонниками трёхкомпонентной теории цвета, основанной на идеях Ломоносова и Ньютона, и сторонниками оппонентной теории, велись жаркие споры. К концу ХХ века эти теории стали считать взаимно дополняющими интерпретациями. В частности Крисс, в своей «зонной теории», предложенной им ещё в начале ХХ века, сделал попытку объединения конкурирующих концепций. Трехкомпонентная теория более была применена для описания функционирования уровня рецепторов, а оппонентная теория — для описания нейронных систем более высокого уровня зрительной системы.
Теория Лэдда-Франклина[]
Предполагается существование сложной фоточувствительной молекулы, которая по-разному реагирует на красный, зеленый, синий и желтый свет, высвобождая вещества, стимулирующие соответствующие нервные окончания. Теория эволюционно ориентирована: двухцветная система зрения объясняется существованием менее высоко развитой молекулы, а ахроматическое зрение — еще более примитивной. Эта теория в настоящее время имеет лишь историческое значение.
Модель цветного зрения Рэнда[]
Модель цветного зрения Эдвина Рэнда (Лэнд??)(Ретинальная теория) предполагает существование трех отдельных зрительных систем (ретинексов), каждая из которых чувствительна преимущественно к одному из световых диапазонов (длинные, средние, короткие волны). Подсистемы функционируют как аналог монохромного изображения, рассматриваемого через определенный фильтр. Каждая подсистема — ретинекс проявляет максимальную активность в ответ на красный, зеленый и синий цвета.
Модель Хартлайна[]
В 1938 году Хартлайном было введено понятие «рецептивного поля». Под рецептивным полем ганглиозной клетки подразумевается тот участок сетчатки, при раздражении которого в конечном итоге меняется частота разрядов данной ганглиозной клетки. Как известно, в сетчатке проявляется довольно четко выраженное латеральное торможение, которое на уровне биполярных клеток осуществляется горизонтальными, а на уровне ганглиозных клеток — амакриновыми клетками. Следовательно при воздействии света на рецепторы к ганглиозной клетке из разных точек сетчатки должны поступать не только возбуждающие влияния, но также и тормозящие. Совокупность этих воздействий, в свою очередь, будет определять функциональную организацию рецептивного поля ганглиозной клетки. Концентрические рецептивные поля состоят из круглой центральной возбуждающей зоны, которая окружена со всех сторон тормозной периферией. В этом случае деление клеток на типы ведется с учетом характера их реакций на раздражение различных зон рецептивного поля. Нейроны возбуждающиеся при освещении центральной зоны рецептивного поля относятся к on — нейронам, а возбуждающиеся затемнением центральной зоны к off — нейронам. В тоже время on — нейрон возбуждается при затемнении периферии, а off — нейрон при ее освещении.
Размеры рецептивных полей ганглиозных клеток существенно различается у разных видов животных. При этом считается, что с размерами рецептивных полей связана острота зрения животного — чем уже рецептивное поле, тем более мелкие детали изображения может различить зрительная система. Этот вывод подкрепляется данными измерений размеров рецептивных полей ганглиозных клеток, связанных с центральными и периферическими участками сетчатки.
Среди других свойств нейронов, связанных с организацией их рецептивных полей, следует отметить избирательность к направлению движения видимых объектов. Такие клетки дают максимальные разряды, когда стимул движется через рецептивное поле в строго определенном направлении, которое таким образом, оказывается предпочитаемым для данного нейрона. Ганглиозные клетки сетчатки обладающие избирательностью к направлению движения, изучены в сетчатках многих видов млекопитающих, в том числе и в сетчатке кошки.Были также предприняты попытки обнаружить корреляцию между типом нейрона и особенностью его спектральной чувствительности. Однако результаты авторов, ведущих исследования в этом направлении, весьма противоречивы. Одни находят, что имеется корреляция между скоростью проведения возбуждения в аксонах ганглиозных клеток и чувствительностью этих клеток к свету с разной длиной волны только для on -нейронов, другие же авторы, наоборот считают, что частота разрядов on -нейронов зависит от интенсивности света, а не от длины его волны, on — off -нейроны же реагируют исключительно на свет.В опросе о конвергенции импульсации на уровне ганглиозных клеток мнения исследователей также расходятся. Имеются данные, согласно которым сигналы от колбочек и палочек в высшие отделы центральной нервной системы поступают по разным путям. Если это так, то можно заключить, что, несмотря на то, что сигналы от рецепторов обоих типов конвергируют на уровне ганглиозных клеток, они все же остаются независимыми друг от друга[16]
Гипотеза Г. Хартриджа[]
В 1947 году появилась «полихроматическая» гипотеза Г. Хартриджа. Он полагал, что помимо трёх основных, первичных рецепторов (оранжевого, зелёного и сине-зелёного) должно быть ещё четыре или пять других дополнительных, или вторичных, включая жёлтую и синюю пару, действующую как единое целое. Модель Г. Хартриджа охватывала практически всю гамму существующих цветов. Однако к этому времени морфология, структура сетчатки и колбочек были уже достаточно хорошо изучены. В сетчатке не обнаруживалось даже двух разных типов колбочек, не говоря уже о семи. В практике же полихроматическая модель давно используется, например в семицветной печати.
Концепция М. С. Смирнова[]
В 1955 году известный исследователь цветового зрения, советский ученый М. С.Смирнов выдвинул новое предположение: все три типа приёмников находятся в одной колбочке. Данное предположение было во времена, когда не возможно было проверить гипотезу на гистологическом уровне с применением приборов (например, флюоресцентных наноскопов) с разрешающей способностью 1‒10нм, позволяющих рассмотреть работу живых экстерорецепторов колбочек и палочек в цвете, в режиме стеро-3D на атомно-молекулярном уровне. Концептия М. С. Смирнова с точки зрния гипотеоической подтвердилась экспериментальными данными (см. Лаборатория Р.Е.Марка). Были замечены при рентгеноскопии среза сетчатки и сетчатки в плане, когда одна колбочка способна в различных ситуациях освещения пучком лучей света предметной точки выделять сигналы трёх основных лучей света RGB, что послужило открытием принципа трихроматизма — основы трёхкомпонентной теории цветного зрения.
Как позже (1975) нелинейная теория цветного зрения С.Ременко в отличие от теории М. С. Смирнова не нашла признания в мире на фоне открытий, связанных с принципом трихроматизма, подтверждённых экспериментально..[17] До настоящего времени сторонники С.Ременко продолжают бездоказательно утверждать, что в цветном зрении участвует блок (палочка+колбочка). Так в 2011 году при специальном исследовании работы палочек и колбочек у рыб, получены графики ретиномоторной реакции фоторецепторов палочек и колбочек, где точно зафиксирована работа палочек и колбочек. При дневном освещении — при цветном зрении работают только колбочки. Палочки работают при сумеречном и ночном освещении в области бело-чёрного цвета. (См. Участник:Миг/Ретиномоторная реакция фоторецепторов).
Теория П. Уолравена[]
Голландский ученый П. Уолравен (иначе его фамилию упоминают, как Валравен) на свей модели практически заложил основы трёхкомпонентной теории цветного зрения. В свей теории он предположил, что в сетчатке человека должны присутствовать три типа колбочек, причём сигналы «красной» и «зелёной» колбочек делятся на три, а «синей» — на две части. Одна часть сигналов трёх колбочек поступает на суммирующий узел, образуя яркостный сигнал. По одной части сигнала «красной» и «зелёной» колбочек подается на второй сумматор, на выходе которого получается жёлтый сигнал. Теперь имеются четыре сигнала: красный, зелёный, жёлтый и синий. Из них образуются два сигнала двух противоположных пар: красно-зелёной и жёлто-синей. Как мы видим, это по сути основа оппонентной теории цветного зрения Геринга. Эту модель можно было бы назвать «телевизионной» — так как она в общих чертах копирует механизм формирования цветовых сигналов в телевидении. Модель П. Уолравена, в общих чертах увязала четырёх- и трёхкомпонентную гипотезы цветного зрения. Позже эту же модель цветовосприятия описали Давид Хьюбл (David H.Hubel) и Торстен Вайзел (Torsten N.Wiesel), (получившие Нобелевскую премию 1981 года за работы, касающиеся принципов переработки информации в нейронных структурах и механизмов деятельности головного мозга). Они предположили, что мозг получает информацию вовсе не о красном (R), зелёном (G) и синем (B) цветах (теории цветаЮнга-Гельмгольца,), а о разнице яркости белого (Yмах) и черного (Yмин), разнице зелёного и красного цветов (G-R), разнице и синего и жёлтого цветов (B-yellow), при этом, жёлтый цвет (yellow=R+G) есть сумма красного и зелёного цветов, а R, G и B — яркости цветовых составляющих — красного, зелёного, и синего.
Получаем систему уравнений — Кч-б=Yмах-Yмин; Кgr=G-R; Кbrg=B-R-G, где Кч-б, Кgr, Кbrg — функции коэффициентов баланса белого для любого освещения. Но эти предположения не могли объяснить механизм цветовосприятия, они касались только способа передачи сигналов от рецепторов в мозг. В этой теории всю обработку информации также относили исключительно к работе головного мозга.
Теория стадий (Мюллер, Джадд, Валравен)[]
Работы Мюллера (Müller’s) охватывает четыре основные области:
психофизика (psychophysics),
память,
психология мышления,
теория цвета.
Психофизика основывается на теории цвета, которая основана на Hering. Работа памяти вводит в психологию мысли, которая основана на Herbart и Külpe.[18]
В 1975 году появилась нелинейная теория зрения советского учёного С.Ременко, предполагающая наличие в сетчатке глаза человека только двух типов фоторецепторов — (палочка+колбочка), обеспечивающих цветное зрение.
В отличии от всех остальных существующих на сегодня теорий она единственная, которая объясняет механизмы обработки сигналов фоторецепторами, поддержание баланса белого цвета и моделирует работу глаза в целом. Она базируется на методах исследований на базе модели глаза (колориметра) c оценкой выходных сигналов на рецепторном уровне, на чисто физических принципах (без учёта субъективных факторов, работы мозга), без учёта третьего фоторецептора ipRGC ганглиозного слоя сетчатки, ретиномоторной реакции фоторецепторов.[19][20][21][22]
В этой связи теория С.Ременко в своём главном выводе не совпадает с данными современных исследований. Так в 2011 году данные иследовний ретиномоторной реакции фоторецепторов на основе гистогенетических исследований на живой клетке на уровне флюоресцентной микроскопии показали, что колбочки и палочки работают изолированно друг от друга с границей раздела — освещённости в 498нм. До 498нм, в условиях сумеречного освещения работают только палочки, а свыше 498нм — колбочки. Это же доказал доктор М. А. Островский, Институт биохимической физики РАН им. Н. М. Эмануэля, Москва в работе[23] (См. также Биохимия зрения и свободно-радикальное окисление) и более фундаментально вопросы цветного зрения исследовал и обобщил учёный Р. Е. Марк. (См. Труды доктора Р.Е.Марка и его лаборатории, Центральная ямка сетчатки глаза, Ретиномоторная реакция фоторецепторов сетчатки глаза). Не случайно нелинейная теория зрения не признана в мире.
Теория цветного зрения Лэнда была опубликована в 1977 г.[24][25]
Лэнд предложил новую теорию цветового зрения, суть которой можно свести к тому, что цвет — это свойство глаза воспринимать любые электромагнитные колебания видимого спектра на двух уровнях — рецепторном и нейронном. На рецепторном уровне восприняты результирующие, интерференцирующие чёрно-белые лучи света разной яркости (амплитуд и длин электромагнитных колебаний), которые на нейронном уровне оппонентно отобраны и отражены в зрительном отделе мозга в виде цветовых ощущений (можно сказать, что это пример метамерии). То есть это как результат действия «длинных волн против коротких». По выражению Лэнда, цвет, как его видит глаз, есть информация «о распределении коротких и длинных световых волн по полю зрения».
Сферическая модель цветового зрения (Соколов, Измайлов)[]
Первый вариант сферической модели цветового зрения, изложенный в работах Соколова, Зимачева, Измайлова (1975), Измайлова и Соколова (1978) Фомина, Соколова, Вайткявичуса (1979), (Sokolov, et al, 1979) то есть, в работах до 1980 года, основывался на экспериментах по различению равноярких цветов, и в качестве основной меры цветового различия (метрики цветового пространства) было принято расстояние по малой дуге большого круга сферы, то есть метрика на сфере единичного радиуса, соответствующая частному случаю римановой геометрии (двумерному пространству с положительной кривизной). Аналогичное положение принято в сферической модели Воса и Варлавена (Vos, Warlavn, 1972).
Специальные экспериментальные исследования метрики цветовых различий, результаты которых изложены в монографии Измайлова (1980), и в последующих работах (Измайлов, 1981, Izmailov, 1982, Sokolov, Izmailov, 1983) заставили отказаться от этого положения, и привели к выводу, что цветовые различия точнее выражаются евклидовой метрикой, а не римановой. Хотя цвета располагаются на сфере, и геодезическая линия смешения цветов задается большим кругом сферы, но цветовое различие как кратчайшее расстояние между двумя точками измеряется не по дуге, а по хорде, то есть, в евклидовой метрике. В этом смысле цветовая сфера это не Риманово (частное) пространство, а представляет собой гиперсферу в евклидовом пространстве.
Этот аспект очень важен, поскольку он позволяет понять и объяснить одну проблему, возникшую при измерении цветовых различий, и сформулированную в работах Мак-Адама (1964) и Джадда (1967), как проблема неаддитивности цветовых различий. Проблема состоит в том, что для любых трех цветовых точек (i, j, k), расположенных в цветовом пространстве на одной геодезической линии сумма различий (D ij + D ik) всегда больше, чем различие Dik. Причем, это расхождение ε = [(D ij + D ik) — D ik] меняется и зависит от величины D. При пороговых различиях (и близких к пороговым) это расхождение минимально и сравнимо с ошибкой измерения. Именно поэтому цветовые функции, основанные на пороговых измерениях, оказались согласованными с цветовыми функциями, выведенными исходя из римановой метрики сферической модели в работе Измайлова и Соколова (1978). Но при переходе к сверхпороговым различиям это расхождение начинает увеличиваться и риманова геометрия уже не годится для модели цветового пространства.
То, что мы видим (точнее, ощущаем) как цвет, представляет собой результат комбинированного действия на глаз и мозг:
1) спектрального распределения световых потокот от источников света;
2) физических свойств всех материалов, пропускающих или отражающих световой поток (особенно часть светового потока, ориентированную в сторону глаза);
3) физиологических реакций зрительной системы на действие светового потока, включающей в себя результаты обработки нейронными сетями нервных импульсов, передаваемых в зрительный центр мозга;
4) переработки нашим мозгом этих сигналов в сочетании с сигналами из соседних областей поля зрения, нашими воспоминаниями о сходных ситуациях, имевших место в прошлом опыте..[26] Это достаточно общее высказывание в целом верно, но для того, чтобы «теория цвета» стала продуктивной, требуется развить и углубить, связать с фактами модель каждого из уровней цветовосприятия.
Теория цветного зрения может считаться полезной только при выполнении следующих условий:
теория должна строиться на объективных экспериментальных данных:
данных с использованием результатлв микроскопии и гистологического анализа (например, люминесцентной микроскопии живых тканей),
учёт работы фоторецепторов колбочек, с точки зрения влияния конической формы внешней доли мембраны, работы её как волновода конической формы в жидкой среде, наличия классов колбочек, меняющие свои размеры, и характеристики, связанные с градиентом уменьшения диаметра большего сечения конуса с одновременным увеличением длины мембраны с центробежным вектором от центра жёлтого пятна к периферии сетчатки. (Феномен, открытый учёным Джон А. Медеиросом, см. Специфика работы мембраны колбочек),
учет различных симптомов всевозможных видов цветовой слепоты,
основываться на биофизических и биохимические законах,
описываться математическими зависимостями в трёхмерном 3D пространстве.[27]
В настоящее время удалось обнаружить, что различий между фоторецепторами сетчатки глаза колбочками нет, однвко принято, что колбочка работает с оппонентным отбором трёх основных цветов RGB в зависимости от падающего на неё луча света предметной точки. Откуда принято считать о наличии трёх типов колбочек. (синей, зелёной, красной).[28] Однако, цветные снимки срезов сетчатки показывают, что любая колбочка работает в зоне частей спектров S,M,L в виде блока, например, у примата RGB — Trichromacy.[29]
В течении более чем сотни лет, на почве множества различных предположений и гипотез была сформулирована Теория многокомпонентного цветного зрения. В основу этой теории вошли предположения М. В. Ломоносова, Томаса Юнга, Гельмгольца, Уолравена. Все они считали, что в сетчатке глаза должно существовать три типа приёмников, чувствительных к узким частям спектра. Только в настоящее время она доказана и получила подтверждения. Большая часть исследователей в настоящее время демонстрируют её. Данные получаемые в различных лабораториях на основе исследований живой клетки уже на снимках демонстрируют в цвете, что только колбочки участвуют в цветном зрении, которые на основе фотопигмента кон-опсины отбирают оппонентно основные лучи спектра в номинальных точках спектра S,M,L. Исследования 2011 г. вопросов ретиномоторной реакции фоторецепторов на срезах живой сетчатки показали, что в условиях дневного освещения, при цветном зрении работают только колбочки. Палочки работают в условиях сумеречно-ночного освещения с чётно-белым видением. При этом колбочки и палочки работают не зависимо друг от друга. (Cм. Мембраны колбочек и палочек и их функция.
Выводы[]
Рентгеноскопия сетчатки и отделов мозга, проведенная ещё в 1966, повторно в 2009 годах Р. Е. Марком, на атомно-молекулярном уровне подтверждает ранее сформулированные гипотезы о работе сетчатки, где заложены фоторецепторы колбочки, выделющие основные лучи спектра
S,M.,L («синие», «зелёные», «красные»), которые на рецепторном уровне воспринимают, трансдукцируют основные сигналы RGB оптического изображения (контурного, рецепторного) перед передачей их в высшие зрительные отделы (нейронные) головного мозга (см. Труды доктора Р.Е.Марка и его лаборатории). При этом следует подчеркнуть, что структурно колбочки устроены одинаково, но в зависимости от места расположения они имеют разные морфологические размеры, например, в центральной ямке сетчатки они самые маленькие, так как плотно упакованы и при этом, в зоне ямочки диаметром до 1 мм расположены только колбочки красные и зелёные, где нет палочек вообще. Каждая колбочка способна оппонентно выделить один из основных цветовых сигналов S,M.,L, RGB, так как фотопигмент опсин в зависимости от сфокусированного и выделенного основного луча предметной точи способен в результате мутации белка пигмента принимать красную, зелёную и синюю окраску!
Большой интерес представляют последние данные исследований Лаборатории Джона А. Медеироса, где далее углублен и освещён широкий диапазон и большое количество проблем, связанных с человеческим цветным зрением на основе метода использования влияния конической формы доли мембраны колбочек и цилиндрических форм мембраны палочек, работающих как конусный волновод и обычный цилиндрический.
Я (Джон А. Медеирос) надеюсь, что в процессе я пролил некоторый свет на тему, Покрасите Видение: Новое Понимание. В то время как я сделал то, чему я верю — очень сильный случай для несоответствия (по крайней мере) модели с тремя конусами цветного видения как основанный на «красных», «зеленых» и «синих» фотопигментах, уникально изолированных в пределах каждого типа колбочек (То есть колбочки все одинаковые, но способны выделять свои полученные лучи света и нужный пигмент), есть все еще, конечно, очень важная роль для фотопигментов в любой модели цветного видения. Фотопигменты обеспечивают первый шаг в трансдукции света в визуальную сенсацию. Однако, я предлагаю, что роль для фотопигментов вспомогательна для основного спектрального механизма дисперсии сокращения способа волновода. Фотопигменты делают работу поглощения и преобразования света в электрический сигнал и отличительное распределение фотопигментов с различными поглотительными кривыми могла вполне увеличить и улучшить операцию основного механизма, предложенного здесь.(Джон А. Медеирос)[30]
Важное исследование и оценка цветного зрения представлена на базе трудов физика Джеральда К. Хата.[31] В труде Пересмотр прежнего мнения процесса видения он обосновал работу фоторецепторов на исследованиях физического взаимодействия света с внешними долями мембран — фоторецепторов сетчаткиглаза. Это участки сетчатки, где происходит первичное взаимодействие со светом. Дж. Хат вводит современную мысль о важности «nanostructural» элементов, когда свет рассматривается как электромагнитная волна в классической физике, и который проходит через пространственные комплексы сетчатки (названные К.Хатом «nano-антеннами»), фильтруется в виде трёх типов сигналов, соответствующих цветовым диапазонам (RGB). Падающий на сетчатку свет следует рассматривать, как поглощаемый в пространственных структурах между смежными колбочкой и палочкой, а не в виде чистого кванта (подразумевается, что фотоны взаимодействуют в пределах непосредственно тел фоторецепторов).
На основании выданных Джоном А. Медеиросом и К. Хатом данных исследований современного понимания механизма цветного зрения следует:
Колбочки и палочки работают раздельно, притом колбочки — в цвете, палочки в режиме до 498нм (его эксперимент — при 450нм — сумеречного, не цветного зрения;
Начальное взаимодействие света с принимающим волну местом происходит в очень быстрой (фемтосекунда или секунды) структуре времени. Поглощенная энергия в этом случае передаётся через более медленный phononic/solitonic механизм через thylakoid мембрану клетки к ограниченному электронному месту смежного кванта. Этот механизм обеспечивает краткосрочную функцию памяти (memristor) соединения двух мест, который несомненно позволяет интеграцию (суммированию) времени визуальной информации изображения от квантовой области времени до человеческих пропорций нервной системы. Применяя это пространственное понятие антенны к размерам Остерберга пространственной плотности колбочек и палочек на относящейся к сетчатке глаза поверхности, и простые процессы подсчета приложений рецептора под каждым относящимся к сетчатке глаза углом падения лучей, можно показать природу светового (видимого спектра лучей) взаимодействия с этой поверхностью и что важно, процесса trichromicity (трихроматизма).
На основе проведённых фундаментальных исследований академика РАН биохимика М. А. Островского в области работы фотопигмента родопсин в палочках[32] (Биохимия зрения и свободно-радикальное окисление) было также подтверждено, что палочки с фотопигментом родопсином участвуют в зрительном процессе в условиях сумеречного и ночного зрения (чёрно-белого).
В итоге на базе проведенных исследований разных лабораторий и иинститутов было ещё раз доказано и подтверждена правота работы учёных Лаборатории Р. Е. Марка и дефакто в мире, что:
Рис. F. Расположение фоторецепторов в сетчатке глаза
При цветном зрении работают колбочки, но в последовательных нужных режимах: трихроматизма, двухроматизма и монохроматизма в зависимости от места расположения каждого класса колбочек, сечения внешних мембран колбочек, которые имеют наименьшие размеры в силу большой плотности в центре сетчатки глаза и имеют увеличивающиеся размеры в диаметре по мере удаления колбочек от центра с уменьшающейся их плотностью. Размеры длин колбочек наибольшие в центре сетчатки. (См. рис. F, Возможности конической доли внешней мембраны колбочки глаза)
Колбочки работают в условиях дневного освещения и воспринимают лучи света сфокусированных предметных точек изображения и выделяют основные лучи S,M,L, обеспечивая цветное зрение. Палочки работают в условиях слабого и ночного освещения и обеспечивают чёрно-белое зрение. (См. Ретиномоторная реакция фоторецепторов сетчатки глаза).
И нормальное объяснение трихроматизму состоит в том, что у обычного человека сетчатка глаза содержит три разновидности «цветных» фоторецепторов, названных колбочками сетчатки глаза. В зависимости от сфокусированной на них предметной точки изображения они способны воспринимать видимые лучи предметной точки и оппонентно выделять из них три вида основных, базисных самых ярких световых лучей S,M,L, (RGB) на рецепторном уровне не в цвете (сетчатка глаза) с последующей передачей их в мозг — на нейронном уровне, где мы в итоге воспринимаем оптическое изображение в цвете.
См. также[]
Участник:Alexandr/Современные концепции световосприятия и цветного зрения
↑Тонквист Г. Аспекты цвета. Что они значат и как могут быть использованы // Проблема цвета в психологии / Отв. ред. А. А. Митькин, Н. Н. Корж. М.: Наука, 1993. С. 5‒53. (С. 7)
↑С. Ременко, «Цвет и зрение», «Картеа Молдовеняскэ», Кишинёв, 1982 г.
↑С. Ременко, «Нелинейная модель измерения цвета и уточнение терминов колориметрии», Всеакадемический семинар по проблемам стандартизации и метрологии, Ташкент, 20 — 25 ноября 1986 год, стр 41 — 42.
↑С. Ременко, «Определение основных понятий в области колориметрии и измерения цветовых параметров излучения», V Всеакадемический семинар по проблемам стандартизации и метрологии Ереван, 16 — 20 ноября 1987 год, стр 58 — 59.
↑С. Ременко, «Как построить глаз», журнал «Техника Молодёжи», 1981 год,№ 7, стр. 21 — 25.
↑Тонквист Г. Аспекты цвета. Что они значат и как могут быть использованы // Проблема цвета в психологии / Отв. ред. А. А. Митькин, Н. Н. Корж. М.: Наука, 1993. С. 5‒53. (С. 7)
Голдсмит Т. Физиологическая основа цветового зрения пчелы // Теория связи в сенсорных системах / Под ред. Г. Д. Смирнова. М.: Мир, 1964. С. 433—448.
Джадд Д., Вышецки Г. Цвет в науке и технике. М.: Мир, 1978. 592 с.
Измайлов Ч. А., Соколов Е. Н., Черноризов А. М. Психофизиология цветового зрения. М.: Изд-во Моск. ун-та, 1989. 206 с.
Крылова А. Л., Черноризов А. М. Зрительный анализатор. М.: Изд-во Моск. ун-та, 1987. С. 6—41, 54—58, 72—76.
Латанов А. В., Леонова А. Ю., Евтихин Д. В., Соколов Е. Н. Сравнительная нейробиология цветового зрения человека и животных // Журн. высш. нервн. деят. им. И. П. Павлова. 1997. Т.47. Вып.2. С. 308—320.
Пэдхем Ч., Сондерс Дж. Восприятие света и цвета. М.: Мир, 1978. 255 с.
Соколов Е. Н., Измайлов Ч. А. Цветовое зрение. М.: Изд-во Моск. ун-та, 1984. 175 с.
Супин А. Я. Нейрофизиология зрения млекопитающих. М.: Наука, 1981. 252 с.
Хьюбел Д. Глаз, мозг и зрение. М.: Мир, 1990. С. 167—197.
Канаев И. И. Очерки из истории проблемы физиологии цветового зрения от античности до ХХ века. Л.: Наука, 1971. 160 с.
Константинов А. И., Соколов В. А., Быков К. А. Основы сравнительной физиологии сенсорных систем. Л.: Изд-во Ленингр. ун-та, 1980. С. 220—237.
Орлов О. Ю. Об эволюции цветного зрения у позвоночных // Проблемы эволюции зрительной системы позвоночных: В 2 т. Новосибирск: Наука, 1972. Т. 2. С. 69—94.
Сурина М. О. Цвет и символ в искусстве // Ростов-на-Дону: ИЦ «Комплекс», 1998. 255 с.
Черноризов А. М. Нейронные механизмы цветового зрения. Автореф. дисс. … д-ра психол. наук. М., 1999. 69 с.
Шеперд Г. Нейробиология: В 2 т. М.: Мир, 1987. Т.1. С. 419—450.
Соколов Е. Н., Зимачев М. М., Измайлов Ч. А. Геометрическая модель субъективного пространства цветовых стимулов. Эргономика. Труды ВНИТЭ, 1975, № 9, с. 101‒122.
Измайлов Ч. А., Соколов Е. Н. Метрические характеристики сферической модели пветоразличения // Вестн. МГУ. Сер. 14. Психология. 1978. № 2. С. 47—61.
Фомин С. В., Соколов Е. Н., Вайткявичус Г. Г. Искусственные органы чувств. М., Наука, 1979.
Sokolov, E. N.; Izmailov, Ch. A.; Izmailova, T. V.; Zimachev, M. M. A spherical model of color vision. Soviet Psychology; 1979. Springer, vol. 17(3), 85‒96.
Измайлов Ч. А. Сферическая модель цветоразличения. М., Изд-во МГУ, 1980.
Измайлов Ч. А. Многомерное шкалирование ахроматической составляющей цвета // Нормативные и дескриптивные модели принятия решений: По мат-лам советско-американского семинара / Ред. колл. Б. Ф. Ломов и др. М.: Наука, 1981. С. 98—110.
Izmailov Ch. A Uniform Color Space and Multidimensional scaling {MDS} In: Psychophysical Judgement and the Process of Perception. 1982 H.G.Geissler and F.Petsold (eds). Berlin,VEB Deutcher Verlag der Wissenschaften, pp.52‒62.
Sokolov E.N. Izmailov, Ch. A The Conceptual Reflex Arc: A Model of Neural Processing as Developed for Color Vision.In: Modern Issues of Perception. 1983, H.G.Geissler, ed., Berlin, VEB Deutscher Verlag der Wissenschaften, р.192‒216
ГР Jacobs, Neitz М., Deegan JF, и Neitz J. Trichromatic окрашивают видение в Новых Мировых обезьянах. Природа 382: 156†"158, 1996. [Medline]
Neitz М. и Neitz J. Молекулярная генетика цветного видения и цветных дефектов видения. Арч Офталмол 118: 691†"700, 2000. [FreeВ FullВ Текст]
Wassle H, Puller C, Muller F, Haverkamp S (2009) Cone contacts, mosaics, and territories of bipolar cells in the mouse retina. J Neurosci 29: 106—117.
Реган до н.э, Jullio C, Simmen B, Vienot F, Чарльз-Dominique P, и Mollon JD. Фрукты, листва и развитие примата окрашивают видение. Фил Транс Р Soc Lond B Biol Наука 356: 229†"283, 2001. [Abstract/FreeВ FullВ Текст]
Реган до н.э, Jullio C, Simmen B, Vienot F, Чарльз-Dominique P, и Mollon JD. Фрукты, листва и развитие примата окрашивают видение. Фил Транс Р Soc Lond B Biol Наука 356: 229†"283, 2001. [Abstract/FreeВ FullВ Текст]
Реган до н.э, Jullio C, Simmen B, Vienot F, Чарльз-Dominique P, и Mollon JD. Фрукты, листва и развитие примата окрашивают видение. Фил Транс Р Soc Lond B Biol Наука 356: 229†"283, 2001. [Abstract/FreeВ FullВ Текст]
Dominy NJ and Lucas PW. Ecological importance of trichromatic vision in primates. Nature 410: 363‒366, 2001.[Medline]
Nathans J. Развитие и физиология человеческого цветного видения: понимания от молекулярных генетических исследований визуальных пигментов. Нейрон 24: 299†"312, 1999. [Сеть Науки][Medline]
Реган до н.э, Jullio C, Simmen B, Vienot F, Чарльз-Dominique P, и Mollon JD. Фрукты, листва и развитие примата окрашивают видение. Фил Транс Р Soc Lond B Biol Наука 356: 229†"283, 2001. [Abstract/FreeВ FullВ Текст]
Neitz M and Neitz J. Molecular genetics of color vision and color vision defects. Arch Ophthalmol 118: 691‒700, 2000.[Free Full Text]
Shapley R. Visual sensitivity and parallel retinocortical channels. Ann Rev Psychol 41: 635‒658, 1990.[Web of Science][Medline]
Dacey DM. Parallel pathways for spectral coding in primate retina. Ann Rev Neurosci 23: 743‒775, 2000.[Web of Science][Medline]
Martin PR. Colour processing in the primate retina: recent progress. J Physiol 513: 631‒638, 1998.[Abstract/Free Full Text]
Dacey DM. Parallel pathways for spectral coding in primate retina. Ann Rev Neurosci 23: 743‒775, 2000.[Web of Science][Medline]
Dacey DM. Parallel pathways for spectral coding in primate retina. Ann Rev Neurosci 23: 743‒775, 2000.[Web of Science][Medline]
Martin PR. Colour processing in the primate retina: recent progress. J Physiol 513: 631‒638, 1998.[Abstract/Free Full Text]
Mullen KT and Kingdom FAA. Losses in peripheral colour sensitivity predicted from «hit and miss» post-receptoral cone connections. Vision Res 36: 1995‒2000, 1996.[Web of Science][Medline]
Calkins DJ and Sterling P. Evidence that circuits for spatial and color vision segregate at the first retinal synapse. Neuron 24: 313‒321, 1999.[Web of Science][Medline]
Martin PR, Lee BB, White AJR, Solomon SG, and Ruttiger L. Chromatic sensitivity of ganglion cells in the peripheral primate retina. Nature 410: 933‒936, 2001.[Medline]
Mollon JD. «Tho' она kneel’d в том месте, где они выращивали …»: использования и происхождение примата красят видение. J Exp Biol 146: 21†"38, 1989.
Silveira LCL, Защита BB, Yamada ES, Kremers J, и DM Охоты. Механизмы Post-receptoral цветового видения в новых мировых приматах. Видение Res 38: 3329†"3337, 1998. [Сеть Науки][Medline]





{kind=link}
{kind=link}
{kind=link}
{kind=link}