{kind=link}
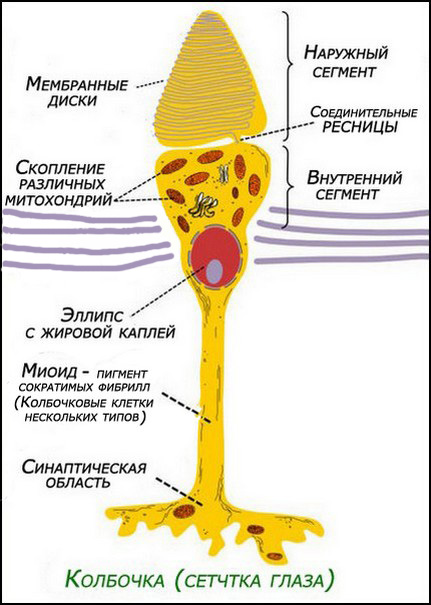
рис. 1. Колбочки (сетчатка)

{kind=link}
рис. 2. Анатомия строения палочки (сетчатка)[1]
Мембраны колбочек и палочек и их функция состоит в восприятии пучков лучей предметной точки изображения c последующим оппонентным отбором основных лучей RGB, выработкой биосигнала (не в цвете, на рецепторном уровне) для передачи его в зрительные отделы головного мозга.
Введение[]
Фоточувствительные фоторорецепторы сетчатки колбочки и палочки — экстерорецепторы, расположенные в сетчатке глаза воспринимают электромагнитное излучение в видимом диапазоне длин волн (рецепторный уровень). Фоторецепторы обеспечивают так-же восприятие оптического изображения (не цветное) — функцию зрения. После передачи сигналов оптического изображения в мозг — в зрительных отделах головного мозга формируется биполярно трёхмерноне, оптическое стереоизображение в цвете (нейронный уровень). Именно тот момент, когда происходит цветное зрение. Оно субъективное и у каждого из нас своё. Мы не можем судит о нашем цвете посредством колориметрии. Колориметр выдаёт цвет, который мы оцениваем у себя по своему, воспринимая отражённые сигналы цветного луча при колориметрии. Например, в работах С.Ременко и его последователей любые цветовые и не только цветовые сигналы электромагнитных волн выдаются фототранзисторами на колориметр, который показывает цвет для нашей зрительной системы. Как можно говорить о нашем зрении, например, цветном не зная где и как расположены палочки и колбочки, как работают фотопигменты в мембранах колбочек. Например, один фототранзистор вообще может заменить все экстерорецепторы сетчатки глаза. Как можно строить графики чувствительности колбочек, палочек, говорить об их связях с новым фоторецептором ганглиозного слоя сетчатки ipRGC не видя их. Можно ли вообще, вообще, писать о цвете, проводя исследования без участия зрительных отделов головного мозга. Больше того, разве можно говорить о работе колбочек и палочек при цветном зрении, отбрасывая данные последнего достижения 2011 г. в области ретиномоторной реакции фоторецепторов. Отсюда давно была отброшена неперспективная концепция С.Ременко.
Согласно ретиномоторной реакции фоторецепторов — механические процессы в сетчатке глаза связанны с перестройкой взаимного расположения рецепторов (палочек и колбочек), и гранул меланина,
в соответствии с уровнем освещённости. Не случайно основной элемент колбочек и палочек мембраны имеют:
- у колбочек — коническую форму,
- у палочек — цилиндрическую.
Колбочки воспринимают свет и выделяют основные лучи света (цвета) RGB с разными длинами волн фронтом поперечного сечения от 8 до 1,5мкм, то под эти лучи размеры в трёх сечениях конуса мембраны имеют приблизительно такие же размеры (см. рис.1). Т.к. при цветном зрении участвуют только колбочки, то они воспринимают и оппонентно выделяют нормализованные лучи спектра S,M.,L (красные, зелёные, синие). Палочки, работая при сумеречном и ночном освещении, воспринимают более сильные синие и ультрафиолетовые лучи, которые в поперечном сечении фронта волны имеют размер 1,5-2мкм, что равно приблизительно поперечному сечению цилиндра мембраны палочек (см. рис.2). Т.е палочки в цветном зрении не участвуют (работают практически в чёрно-белом зрении с элементами сине-голубого оттенка).
Фотопигменты в мембранах колбочек и палочек[]
Зрительные пигменты находятся в бислойных биомембранах, во внешней доле фоторецептора (в мембране колбочек, палочек). Зрительные пигменты и передача зрительного сигнала (визуальная трансдукция) — это комплекс понятий для описания фототрансформации пигментов и их регенерации, процессов передачи сигнала, происходящих в глазу позвоночных животных. Эти биохимические процессы проходят при воздействии света с различной длиной волны (разного цвета) связанные с изменениями в структуре и взаимодействиях зрительных пигментов.
Опсины в мембранах колбочек и палочек[]

{kind=link}
Рис.1.[2]

{kind=link}
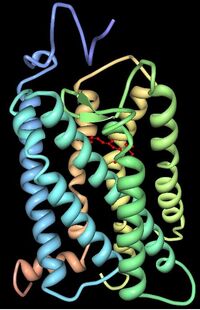
Трехмерная структура бычьего родопсина. Семь трансмембранных доменов показаны разными цветами. Хромофор показан красным цветом.
Образование оптического изображение при зрении у позвоночных животных связано с находящимися к сетчатке глаза колбочек и палочек. Палочки работают при очень слабых уровнях освещения т. н. сумеречном и ночном зрении. Они высоко чувствительны и могут сигнализировать о поглощении одиночных фотонов. Колбочки намного менее чувствительны к свету, чем палочки. Они участвуют в цветном зрении при достаточно ярком (дневном) свете (см. рис. 1).
- На рис. 1. светлопольные изображения фоторецепторов (палочки и колбочки), выделеных из сетчатки саламандры. Фототрансдукция имеет место во внешней доле, в то время как митохондрии плотно упакованы в эллипсоиде.
Фотопигменты опсины и другие зрительные пигменты найденны в клетках фоторецепторов (в мембране) в сетчатках глаз. Опсин образован в виде связки из семи трансмембранных альфа-спиралей, связанных шестью петлями. В клетках палочек молекулы опсинов — род-опсинов) вложены в мембраны дисков, которые размещаются полностью в мембране. «Голова» N-конечной-остановки молекулы простирается внутрь диска, а «хвост» с C-радикалом простирается в цитоплазме клетки. В клетках колбочки (в наружной части мембраны) c кон-опсинами)[3] диски определены плазменной мембраной клетки так, что голова N-конечной-остановки простирается вне клетки. Опсины сетчатки глаза ковалентно связаны с лизином на трансмембранной спирали, самой близкой C-конечной-остановки белка через Шиффовы основания. Формирование изменений Шиффова основания вовлекает удаление атома кислорода от относящегося к сетчатке глаза и двух водородных атомов от свободной группы аминокислоты лизина, давая H2O. Ретинилиден — двухосновная группа, сформированная так, что удаляет атом кислорода относящегося к сетчатке глаза, и потому опсины назвали ретинилиденовыми (retinylidene) белками.
- Классификация опсинов.
| Ретиналь | Тип опсина | Пигмент | Длина волны света с макс. поглощением λMax, nm | Beispiele für das Vorkommen |
|---|---|---|---|---|
| 11-cis-Retinal | Скотопсин | Родопсин | 500 | Человек, Wirbeltiere, Gliederfüßer, Weichtiere |
| 11-cis-Retinal | UV-Фотопсин | UV-Йодопсин | 340 | Honigbiene |
| 11-cis-Retinal | S-Фотопсин | S-Йодопсин | 430 | Affen |
| 11-cis-Retinal | M-Photopsin | M-Iodopsin | 535 | |
| 11-cis-Retinal | L-Photopsin | L-Iodopsin | 565 | Altweltaffen |
| 11-cis-Retinal | XL-Photopsin | XL-Iodopsin | 620 | Vögel |
| 3,4-Dehydro-11-cis-Retinal | Skotopsin | Порфиропсин | 520 | Süßwasserfische, Амфибии |
| 3,4-Dehydro-11-cis-Retinal | UV-Photopsin | UV-Цианопсин | 360 | |
| 3,4-Dehydro-11-cis-Retinal | S-Photopsin | S-Cyanopsin | 420 | |
| 3,4-Dehydro-11-cis-Retinal | M-Photopsin | M-Cyanopsin | 530 | |
| 3,4-Dehydro-11-cis-Retinal | L-Photopsin | L-Cyanopsin | 580 | |
| 3,4-Dehydro-11-cis-Retinal | XL-Photopsin | XL-Cyanopsin | 620 | |
| 9-cis-Retinal | Skotopsin | Iso-Rhodopsin | 485 | |
| 9-cis-Retinal | Photopsin | Iso-Iodopsin | 515 | |
| 3,4-Dehydro-9-cis-retinal | Skotopsin | Iso-Porphyropsin | 510 | |
| 3,4-Dehydro-9-cis-retinal | Photopsin | Iso-Cyanopsin | 575 | |
| 13-cis-Retinal | Bakterien-Opsin | Bakteriorhodopsin, «Halorhodopsin» | 560 | Halobakterien (lichtgetriebene Protonenpumpe, siehe (Chemiosmotische Kopplung) |
| Melanopsin | 485 | Mensch, retinale Ganglienzellen |
Распределение палочек и колбочек в человеческой сетчатке[]
Распределение палочек и колбочек в человеческой сетчатк — необходимая информация для понимания взаимодействия и связей клеток и фоторецепторов в фокальной поверхности сетчатки.
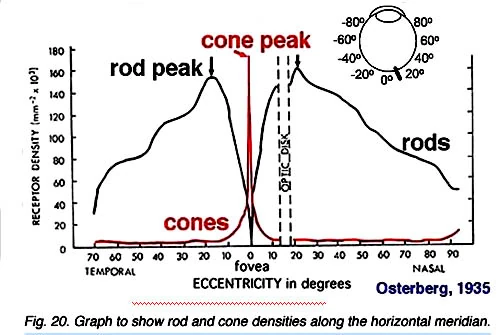
Это для нас очень важно для понимание организации зрительных связей палочек и колбочек в сетчатке. Это связано со знанием пространственного распределения различных типов клеток в сетчатке. Изветно, что фоторецепторы распределены и собраны в блоках с довольно строгой мозаикой. Поскольку мы видели, что в ямке, мозаика расположения колбочек — шестиугольная упаковка. Вне ямки, палочки разбивают близкую шестиугольную упаковку колбочек, но все еще позволяют организованную архитектуру с колбочками, скорее равномерно раздельными и окруженны кольцами палочек. Таким образом, в терминах оценки колбочек и палочек — распределения различных поселений фоторецепторов в человеческой сетчатке. Ясно, что плотность колбочек является самой высокой в foveal яме и падает быстро вне ямки с уменьшением градиента плотности с вектором в сторону движения к периферийной части сетчатки (Osterberg, 1935; Curcio и др., 1987). Есть пик фоторецепторов колбочек в кольце вокруг ямки и равен приблизительно диаметру 4.5 мм или 18 степеням (величина угла конуса) от foveal ямы. Оптический нерв (мертвая точка) — конечно свободный фоторецептор (см. ниже).

{kind=link}
Рис.20. [4]

{kind=link}
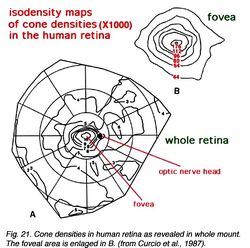
Рис.21. [5]
- Рис.20. Графики распределения палочек и колбочек по горизонтальному меридиану.
- Рис.21. Удельные веса распределения колбочек в человеческой сетчатке.
Важно, что палочек в радиусе круга жёлтого пятна около 3мм вообще нет. Сразу можно сказать, что при дневном освещении, при цветном зрении работают только колбочки. Это доказано при микроскопии среза сетчатки рыб в 2011 году (см. Ретиномоторная реакция фоторецепторов).
Ретиномоторная реакция колбочек и палочек[]
Ретиномоторная реакция фоторецепторов является общим свойством зрительного аппарата как для насекомых[6], так и для позвоночных животных. Особенно активно изучалась ретиномоторная реакция рыб[7]. По данным Наумова и Карташева (1979) у костистых рыб на свету пигментные клетки расширяются и накрывают находящиеся около них палочки. Колбочки продвигаются к ядрам этих клеток и таким образом передвигаются в открытую для прохождения света зону. В темноте же к ядрам подтягиваются палочки и оказываются в зоне ближе к открытой поверхности, где проходит свет. Колбочки в это время приближаются к пигментному слою, а сократившиеся в темноте пигментные клетки прикрывают их[8],[9].
Вследствие ретиномоторной реакции палочки с более высокой светочувствительностью, чем у колбочек, экранируются пигментными гранулами от попадания на них света с наступлением дневного освещения и с наступлением темноты палочки продвигаются в зону с открытым пространством для попадания света, а колбочки экранируются в это время пигментными гранулами и опускаются в зону ниже палочек, куда не проходят более сильные синие и ультрафиолетовые лучи, которые воспринимают палочки. Пиковой зоной раздела длин волн спектра электромагнитных колебаний падающих лучей света на палочки и колбочки является длина волны 498нм. Палочки работают в зоне спектра длин волн синего и ультрафиолетового излучения до 498нм, колбочки работают в зоне более длинных волн после 498нм, за исключением Колбочек-S (синих), пиковая зона которых лежит в зоне длины волны 434нм. Это колбочки расположенные за пределами зоны желтого пятна, окружённые палочками. В любом случае процесс ретиномоторной реакции затрагивает все колбочки и все палочки: все колбочки видят дневной свет, палочки видят ночью и в сумерках только синие и ультрафиолетовые лучи света.
Ультраструктура синаптических окончаний палочки и колбочки[]


{kind=link}
Рис. 23a[10]


{kind=link}
Рис.23b[11]
Работа клеток фоторецептора в сетчатке состоит в трансдукции светового сигнала, т.е. во взаимодействии с полученным квантом света в зрительном процессе, с работой мембраны фоторецептора, содержащей пигмент внешней доли и передачи сообщения , относительно чисел квантов света и фоточувствительности к различным длинам волны, к следующей стадии интеграции (сжатии сигнала при оппонентном отборе) и обработкой его во внешнем сетевидном слое (см. Визуальная фототрансдукция).
Структура синапсов палочек и колбочек[]
Структурное образование, обеспечивающее передачу сенсорной информации от клетки-колбочки именуется "ножка", а от клетки-палочки - "шарик". Ножки колбочек — большие, конические, плоские окончания (диаметром 8-10 мкм) аксонов колбочки, которые лежат более или менее рядом на том же самом узле на внешнем краю внешнего сетевидного слоя (OPL) (Рис. 23a и b). Более многочисленные шарики палочки, напротив, являются маленькими круглыми расширениями аксона (диаметром 3-5 мкм ) или даже расширения тела клетки. Они лежат упакованные между и выше ножки конуса-колбочки (рис. 23a и b). Синаптические окончания обоих типов фоторецепторов заполнены синаптическими пузырьками. В их синапсах к нейронам второго порядка (биполярные и горизонтальные клетки), шарики палочек и ножки колбочек образуют плотные структуры, именуемые синаптическими лентами, указывающие на постсинаптические вставленные процессы (звездочки на рис. 24).
В ножке колбочке приблизительно 30 из этих лент образуются и связаны с 30-ю "триадами" вставленных процессов (Ahnelt и др., 1990). В шарике палочки 2 ленты связаны с 4 вставными нейронами второго порядка, в то время как ножка колбочки поставляет информацию более, чем ста нейронам второго порядка (рис. 23b).

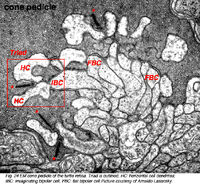
{kind=link}
Рис.24 Ножка колбочки сетчатки черепахи.[12], (фото Арнальдо Ласанский)


{kind=link}
Рис. 25. Шарик палочки черепахи [13]
Колбочка «триады», вставленных процессов второго порядка обычно состоит из центрального элемента, который является древовидным терминалом, вставляющейся биполярной клетки (IBC), и два боковых элемента, которые являются древовидными терминалами горизонтальных клеток (HC) (Рисунки. 24 и 27). Кроме того, другие варианты биполярной клетки имеют дендриты, устанавливающие синаптические контакты на и под поверхностью конуса мембраны колбочки pedicle заключаются в том, что сначала их называли плоскими контактами (FBC) (Missotten, 1965; Dowling и Бойкот, 1966; Kolb, 1970) (рис. 27), но тогда они были лучше охарактеризованы и определены Lasansky (1971) как основные соединения (рис. 26).


{kind=link}
Рис. 26 Ножки колбочки черепахи и типы биполярных синаптических контактов[14]

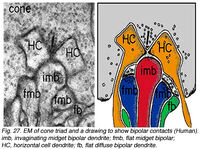
{kind=link}
)Рис.27 Триада колбочки[15]
Шарики палочек имеют только две синаптических ленты, связанные с двумя боковыми элементами, которые являются горизонтальными терминалами аксона клетки (HC) и два центральных вставляющихся дендрита биполярных клеток палочки (rb) (Missotten, 1965; Dowling и Бойкот, 1966; Kolb, 1970). На шариках палочек нет никаких основных соединений.
Это ещё раз показывает, что палочки весьма изолированны от колбочек и других элементов сетчатки (см. рис. 23a, 23b, 24, 25, 26, 27, 28), и формула, что при цветном зрении работают совместно колбочки и палочки, то на уровне показанных структур колбочек и палочек мы лишний раз видим, что это согласуются с вопросами ретиномоторной реакции палочек и колбочек, где палочки при цветном зрении не работают. Приведенная информация снимков фоторецепторов палочек и колбочек на клеточном уровне в цвете — яркое доказательство того, что принцип трихроматизма — не предполагаемый, а реально существующий.

{kind=link}
Рис.28 Триада палочки[16]
См. также[]
Примечания[]
- ↑ Human Physiology and Mechanisms of Disease by Arthur C. Guyton (1992) p.373
- ↑ http://webvision.med.utah.edu/Phototransduction.html#Introduction
- ↑ http://humbio.ru/humbio/ssb/001181d1.htm
- ↑ http://webvision.med.utah.edu/photo2.html
- ↑ http://webvision.med.utah.edu/photo2.html
- ↑ В. П. Тыщенко ФИЗИОЛОГИЯ НАСЕКОМЫХ М., «ВЫСШАЯ ШКОЛА» 1986, с. 223
- ↑ Загальская, Е. О. Морфологические особенности ретиномоторной реакции у молоди симы (oncorhynchus masou) в магнитном поле и красном свете / Е. О. Загальская, В. П. Гнюбкина, А. А. Максимович // Морфология : научно-теоретический медицинский журнал. — 2004. — Том 126,N 6 . — С. 32-36.
- ↑ http://zoometod.com/ixt/ixtiolog_63.html Анисимова И. М., Лавровский В. В. Ихтиология
- ↑ http://clck.yandex.ru/redir/AiuY0DBWFJ4ePaEse6rgeAjgs2pI3DW99KUdgowt9Xt0dmymRAsAFAC41YGK3IYaGvvsyeXdZeHQNpszBsrsqZIe04jRS7Q_-UGVS_eXc43ES6DTijTRmmmHQAhjEN6B9_b0UYDggTd1O0pMdSGPHLJk-QS9esTl7vOPp-wBR0SDO7MxYWW-sQ?data=UlNrNmk5WktYejR0eWJFYk1LdmtxdmdzQ3hIMGVkazluRW9jUlZqeVhkSzNNMXM0amVVYTBXR2h3dFFKZ2xMWDhNY3p5NS1iUUV5aGhjdWlpNFI2OGpRTmlRNEV1ekw5LWpxRm1mYldzWUt2c3VHR1dQVmJYOUlpaVlGUnRmVXdwaGhFaU5ycjIzNA&b64e=2&sign=032480085342e016d3fecd85afeb18ec&keyno=8&l10n=ru&mc=4017&w=862&h=399 Е. О. Загальская, В. П. Гнюбкина. Ультраструктура пигментного эпителия сетчатки глаз молоди симы Oncorhynchus Masou. Биология моря, 2006, том 32, № 1, с. 55-59
- ↑ http://webvision.med.utah.edu/photo2.html
- ↑ http://webvision.med.utah.edu/photo2.html
- ↑ http://webvision.med.utah.edu/photo2.html
- ↑ http://webvision.med.utah.edu/photo2.html
- ↑ http://webvision.med.utah.edu/photo2.html
- ↑ http://webvision.med.utah.edu/photo2.html
- ↑ http://webvision.med.utah.edu/photo2.html