Визуальное цветное зрение — основа современных теорий цветного зрения.
Цветовое зрение, происходящее в зрительной системе инициируется поглощением света с помощью трех различных спектральных классов "шишек" (колбочек). Следовательно, цветовое видение описывается как трёхвариантное восприятие оснвных цветов или как восприятие, ощущение цвета. Первоначально психофизические исследования показали, что цвета могут быть настроены на использование трех различных системах (праймериз). В 1802 году, Томас Молодых предложил модель, по которой восприятие цвета может быть закодировано на три основных цвета фоторецептоов, но не на кодировании тысяч цветовых рецепторов для отдельных цветов.[1]
Спектральная чувствительность колбочек[]

Доктор Kalloniatis теперь Роберт г. Leitl профессор Оптометрии кафедры Оптометрии и Видение Науки, университет Auckland[2]
Спектральная чувствительность колбочек может быть определена посредством нескольких методов. Два из этих метода включают изолирующую рецепторные (receptoral) ответы (Бейлор et al., 1984) с использованием вычисленных от цвета функции нормалей и дихроматизма или двухкомпонентности восприятия цвета (dichromats) (Смит и покорного, 1975; dichromat является предметом, в основе которого сетчатка имеет одну колбочку с фотопигментом в мембране (photopigment), достаточного для этого), микроспекромитрии (microspectrometry) (Bowmaker и Dartnall, 1980) или на основе отражения световых лучей — денситометрии (Раштон, 1963, 1966). В microspectrometer метод предполагает выделение одной колбочки, пропуская свет через неё. Изменения в передаче различных длин волн может быть использовано для вычисления спектрального поглощения колбочкой или возможность определить изменения в электрическом ответе. Отражение денситометрии включает режиссуру, структуру света в сетчатке и определение изменения в поглощении как функция длины волны. Эти результаты впоследствии используются для расчета спектральных поглощений.
Было принято три класса колбочек в человеческой сетчатке глаза, которые были изолированы от указанных технологий. Эти три класса "шишек" представляют:
- Короткие волны чувствительности S-колбочки (S-конуса),
- Средние волны чувствительности M-колбочки (М-конуса),
- Длинноволновую чувствительность L-колбочки (L-конуса).
У всех разная, но пересекающаяся спектральная чувствительность. Спектральная чувствительность S-колбочек с пиком, примерно, в 440 нм, М-коолбочек — 545 нм и L-колбочек с пиком в 565 нм после исправлений, для предварительной потери света сетчаткой. Хотя различные методы измерения дали результаты в несколько различных вариантах максимального значения чувствительности (рис. 1).

Рис.1. Спектральная чувствительность S-колбочек, М-колбочек и L-колбочек. Комбинированные результаты от разных авторов, используя различные способы, в том числе сетчатки денситометрия от Раштон (т и Ñ ), microspectrometry от Коричневого и Вальд (n и “ ) и прирост порог производству искусственного monochromasy от Brinley (D и s ) и прирост порог измерений от Wald (5 ) (От Моисея, р. а., Харт, в. м. (Ред.), Адлер " Физиология Глаза, Клиническое Применение. Сент-Луис: C. V. Мосби Компании, 1987 [3]
Трёхцветная природа цвета по Matches[]
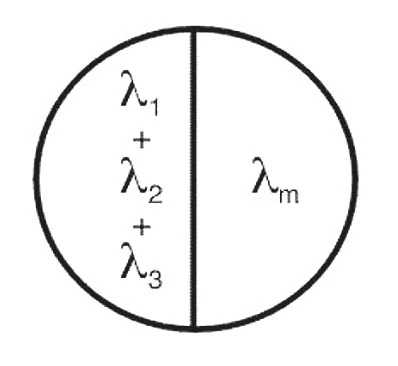
Рис. 2. Для настройки трёх основных лучей цвета на левой стороне двустороннего поля по Матчу стандартный цвет представлен на правой стороне двустороннего поля[4]
Трехцветная природа цветового зрения позволяет практически получить любой цвет, при смешении трех основных цветов (RGB). При этом trichromacy цветного зрения является линейной функцией. Это означает, что колориметрические уравнений имеют свойства обыкновенного линейного уравнения.
Цвет, соответствующий количеству Cs стимула S, может быть выражен как:
Cs(S) = C1(1i) + C2(2i) + C3(3i)
где li-3i есть три основных цвета (не обязательно монохроматического цвета спектра) и C1-C3 - трехстимульные значения.
Трехстимульные значения представляют собой количества каждого из трех первичных выборов, необходимых для достижения соответствия для цвета и яркости (рис. 2). Они могут быть выражены в единицах светового потока или потока излучения, или даже произвольный масштаб, при условии, что произвольный масштаба передает относительные пропорции трёх количеств.
Аддитивный и субтрактивный синтез цветов[]


Рис.3.С трех цветов: синего, зеленого и красного RGB


Рис.4. Субтрактивные цвета смеси голубой, пурпурный и желтый — применяют для получения синего, зеленого и красного цветов.[5]
Голубой, пурпурный, желтый и белый формируются из смесь этих цветов. Два или более цвета могут быть сложены вместе для создания нового цвета, состоящий из смеси первоначального цвета. Это можно продемонстрировать на белом экране с трех цветов: синего, зеленого и красного. Голубой, пурпурный, желтый и белый формируются из смесь этих цветов (рис. 3). Одним из требований в выборе цвета, которые будут использоваться в цветовое смешение экспериментов, является то, что два из них не могут быть смешаны для получения третьего.
Субтрактивные цвета смеси включают выборочно поглощение волн. Голубой, пурпурный и желтый при вычитания цветов (праймериз). Если белый свет, проходящий через желтый фильтр, то желтый фильтр будет поглощать синий и передавать красный и зеленый, что делает желтый цвет. Поэтому, желтый цвет может рассматриваться как a-B фильтр. Пурпурный фильтр вычитает или поглощает зеленый (-G фильтр) от белого света и голубой — вычитает или поглощает красный (-R фильтр) от белого света (рис. 4).
Законы Грассмана обеспечивают количественное описание цвета (Grassman 1855). Они держат хорошо в предписанные набора условий (относительно яркости, адаптации наблюдателя, размера поля, и т.д.). Законы Грассмана полезны для количественного выражения данных цветов, но, как правило, на данные цвета влияют следующим образом:
- Макулярный пигмент (ксантофилла) и вариации связаны с центральным 4 - 5 градусов видением;
- Хроматические аберрации;
- Род заражения, особенно при больших полях и участия низких photopic световых уровней;
- Отказ Закона Эбни яркости аддитивности, в том числе эффекта Гельмгольца-Кольрауша;
- Безольд-Brücke hue переход на яркие интенсивности.
Спецификация цвета[]
Манселл — цветовая модель[]
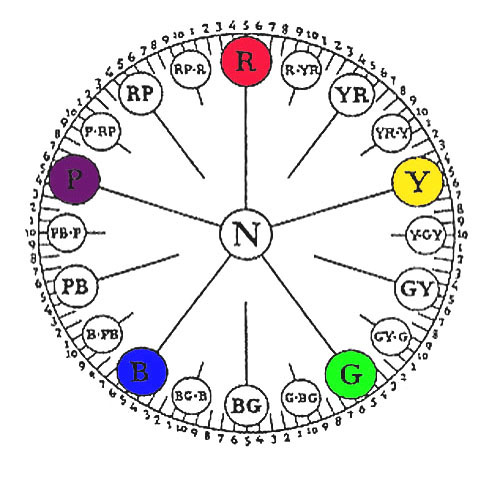
Рис.5. Манселл топ, показывающий расположение различных цветов[6]


Рис.6. Корпус: система цвета, иллюстрирующая hue, величину и цвет.[7]


Рис.6a.Цветовая модель Манселла — оригинал[8]
Эти три измерения соответствуют трем восприятиям, атрибутам человеческого цветового зрения:
- Тон(chroma),
- Насыщенность (цвета)(hue),
- Светлота цвета(Value)
Или определяют три характеритики схем цветовых моделей Манселла: 1.Насыщенность (цвета) — Hue: характеристики, связанные с длинами волн или доминирующей длины волны. Hue обозначается комбинацией букв и цифр, составляющих 100 шагов шкалы (рис. 5). Есть десять разделов категорий, используемых для обозначения hue, с каждым из них далее подразделяется (используя цифры от 1 до 10) на десять подгрупп. Если цифры, обозначающие hue подгруппа 5, то это может быть опущено (например, 5R-это же hue как R).
2.Светлота цвета(Value) — Стоимость (светлота): Стоимость указана на числовой шкале от 1 (черный) до 10 (белый), и это атрибут, связанный с отражательной способностью и светимостью (или светлый).
3. Тон(chroma) — Цветность: Chroma-Munsell характеристика, соответствующая насыщенности. Это указано по номерам, по шкале от 0 до различных величин и зависит от насыщенности участка.
Например, цвет может иметь обозначение 2GY 6/10. Это означает, что он-зеленый/желтый, что является довольно близким к тому, желтому; это имеет значение 6 (т.е. почти на полпути в черный/белой шкале) и цветности 10 (т.е. это насыщенный).
Масштабирования, используемые в корпусе система предназначены для его создания формы. Другими словами, цвета образцов расположены в равных visual шагах. Например, ощущение разницы между chroma 3 и цветностью 4 (номинально) то же, как ощутимая разница между цветностью 4 и 5. Этот масштаб является одинаковым для всех трех измерений, хотя размеры шагов по разным размерам не сопоставимы (то есть, один шаг разница в hue не имеет такого же восприятия разницы в один-единственный шаг в насыщении).
Схематическое представление манселл системы показана на рисунке 6. Значение масштаб по вертикальной оси, hue масштаб по периметру цилиндра и цветности находится на радиальной шкале. Манселл сверху, как показано на Рис. 5, даёт более точное представление об этой концепции.
Рис. 6. Корпус : цветовая система, иллюстрирующая hue, величину и цвет.
В 1931 году Комиссия Internationale dElairage (CIE) приняла систему цветности для RGB (цветовая модель)
Принцип трихроматизма[]
- Принцип трихроматизма включает:
- Идентификация спектральной чувствительности двух пигментов колбочек относящейся к сетчатке глаза денситометрией Руштона (Руштон, 1963).
- Идентификация трех пигментов колбочек микроспектрометрией (Марки, Dobelle и MacNichol, 1964).
- Идентификация генетического кода для красных колбочек — L, зелёных колбочек — М. и синих колбочек — S (Nathans и другие, 1986a, b).
- Цвет, соответствующий функциям.
- Изолируя фоторецепторы и измерение их физиологического repsonses как функция длины волны (Baylor и другие, 1984).
- Спектральные размеры чувствительности (Wald-Marre — фунуционирование спектральной чувствительности и функционирование « Турникетов» механизмов).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Если теория трихроматизма (trichomatic) ранее была не в состоянии составлять четыре уникальных цвета: красный, зеленый, желтый и синий, и также не в состоянии объяснить, почему dichromats может чувствовать белый и желтый, то в настоящее время Трихроматизм с открытием третьего фоторецептора ipRGC сетчатки с фотопигментом меланопсином, с расшифровкой изменчивости опсинов, которые входят в мембраны клеток фоторецепторов палочек и колбочек в виде разновидностей опсина как родопсин, йодопсин и др., позволяют дифференцицироваться колбочкам и реагировать на нормализованные спектральные зоны лучей S,M,L (синих, зелёных, красных), участвуя в оппонентном отборе более ярких сигналов. Недавние открытия ретиномоторной реакции фоторецепторов палочек и колбочек, проведенные флюоресцентные микроскопические исследования сетчаток птиц дали возможность рассмотреть клетки колбочек, палочек на молекулярном уровне в цвете и стерео, где мозаика сетчатки содержит блоки из четырёх колбочек (четырёхроматизм): фиолетовой, синей, зелёной и красной и т.д. Модель оппонентного цветного зрения в сочетании с работой нейронов мозга рассматривается на уровнях рецепторном и нейронном, которые дополняют друг друга. И вообще уже вопрос многокомпонентной модели цветного зрения как трихромаик, четырёхроматик и т.д. не подлежит рассмотрению. (См. Цветное зрение у птицЛаборатория Р.Е.Марка, Ретиномоторная реакция фоторецепторов, Фоточувствительные клетки сетчатки ipRGC, Опсины).
Вопросы функции «расовой дискриминации» и объекты протипоставления цвета противника (красный-зелёный, синий-жёлтый, белый-чёрный) в связи с вышеизложенным разрешаются.
Таким образом, теория трихроматизма рассматривается только в сочетании с теорией цвета противника, предложенная Ю.Герингом в 1872 вначале конкурировала с хорошо принятой trichromatic теорией, которая объясняет trichromaсy видения и предсказывает цветовые зоны чувствительности. Противник Ю.Геринга окрашивает, теория предлагает, что есть три канала: красно-зеленый, синий-желтый и черно-белый, с каждым ответом с оппонентным принципом отбора цвета. Таким образом, или красный или зеленый воспринят и никогда зеленоватый-красный. Ю.Геринг, однако, никогда не бросал вызов начальным стадиям обработки выраженного в соответствии с trichromatic теорией. Он просто утверждал, что любая цветовая теория видения должна объяснить наше восприятие, то есть, цветовое сопротивление как показано цветным после отбора изображений.
Hurvich и Jameson (1957) обеспеченные количественные данные для цветового сопротивления. Используя парадигмы отмены оттенка, были изолированы психофизические цветовые каналы противника. Vl функция использовалась к дискриминации яркости, чтобы описать восприятие черноты и белизны. Поэтому, регулируя количество синих или желтого, красного или зеленого, любая типовая длина волны может быть подобрана (фигура 15). Дополнительные длины волны могут использоваться, чтобы отменить друг друга для всех длин волны кроме четырех уникальных оттенков (синий, зеленый, желтый и красный).


{kind=link}
Рис. 15.[9]
- Рис.15. Hurvich и эксперимент Jameson, используя синий или желтый, красный или зеленый, которые соответствуют всем длинам волны видимого спектра (Hurvich и данные Джамезона (1957) от Бенджамина, W. J. (Эд), Клиническое Преломление Бориша. Филадельфия: W. B. Компания Saunders, 1998).
Другое свидетельство, поддерживающее теорию цвета противника включает:
- Электрическая регистрация горизонтальных ячеек от сетчатки рыбы показывает синий-желтый процесс противника и красно-зеленого противника (Svaetichin, 1956).
- Электрическая регистрация от бокового geniculate ядра, показывая противнику окрашивает процессы (DeValois и другие, 1966).
- Электрическая регистрация ячеек нервного узла от сетчатки примата, показывая противнику окрашивает процессы (Gouras, 1968; de Monasterio и Gouras, 1975; Zrenner и Gouras, 1981).


{kind=link}
Принципиальная схема трёхкомпонентного цветного зрения человека, приматов на примере работы колбочек, палочек, ipRGC, головного мозга и явления ретиномоторной реакции фоторецепторов.


{kind=link}
Принципиальная схема организации цветного зрения (на примере сетчатки цыплёнка).
А. Фоторецептор-колбочка. Свет проходит сквозь хрусталика и до пигментов колбочки расположенных на её «дне».
Б.1-Б.4 У птиц обычно четыре типа колбочек (они «тетрахроматы»), что позволяют им синтезировать многоцветное изображение, и различать цвета лучше, чем человек. Обработка полученных разностных сигналов, происходящая в нейронной сети сетчатки (десятки типов клеток) обеспечивает возможность ясного различения тысяч цветов и оттенков.
Стадия Theory:This привела к современной модели нормального цветового видения, которое включает и trichromatic теорию и теорию цвета противника в две стадии (фигура 16). Первую стадию можно рассмотреть как стадия рецептора, которая состоит из этих трех фотопигментов (синие, зеленые и красные конусы). Второе - нервная стадия обработки, где цветовое сопротивление происходит. Вторая стадия - на post-receptoral уровне, и происходит уже на горизонтальном уровне ячейки (рецепторном, в дисках мембраны перед выдачей сжатого сигнала в мозг).[10]
Выводы[]
В данном исследовании доказан принцип трихроматизма. Т.е. в цветном зрении участвуют только колбочки, выделяющие синие, зелёные, красные сигналы предметных точек изображения, которые оппонентно отбираются дополнительно и отправляются по зрительным нервам в зрительные отделы головного мозга, где формируется оптическое изображение в цвете. Фоторецепторы палочки здесь не участвуют. Выводы нелинейной теории зрения, что при цветном зрении работают блоки палочка+колбочка не правильны. (См. ещё Ретиномоторная реакция фоторецепторов).
См. также[]
Примечания[]
- ↑ http://webvision.med.utah.edu/book/part-viii-gabac-receptors/color-perception/
- ↑ http://webvision.med.utah.edu/book/part-viii-gabac-receptors/color-perception/
- ↑ http://webvision.med.utah.edu/book/part-viii-gabac-receptors/colo perception/
- ↑ http://webvision.med.utah.edu/book/part-viii-gabac-receptors/color-perception/
- ↑ http://webvision.med.utah.edu/book/part-viii-gabac-receptors/color-perception/
- ↑ http://en.wikipedia.org/wiki/HSL_and_HSV
- ↑ http://en.wikipedia.org/wiki/HSL_and_HSV
- ↑ http://en.wikipedia.org/wiki/HSL_and_HSV
- ↑ http://webvision.med.utah.edu/KallColor.html
- ↑ http://webvision.med.utah.edu/KallColor.html