Принцип трихроматизма в цветном зрении принят во всём мире. Согласно нему при цветном зрении участвуют одинаковые колбочки в блоках не менее трёх в блоке, воспринимающие видимые лучи света. При этом идёт оппонентный отбор основных лучей спектра RGB - нормализованных сигналов S,M.,L, формирующих нецветные аналоговые сигналы для отправки в мозг. При этом из всего пучка светового потока предметной точки, спроектированного на любую колбочку блока колбчек, отбираются наиболее яркие лучи света. Сторонники нелинейной теории цветного зрения считают, что цветное зрение осуществляется блоком «колбочка+палочка», и что колбочек, воспринимающих синие (ультрафиолетовые) лучи видимого спектра света не существует.[]
Введение[]
{kind=link}
Рис.R. Синяя колбочка на снимке среза сетчатки при рентгеноскопии по данным Лаборатории Р.Е.Марка
{kind=link}
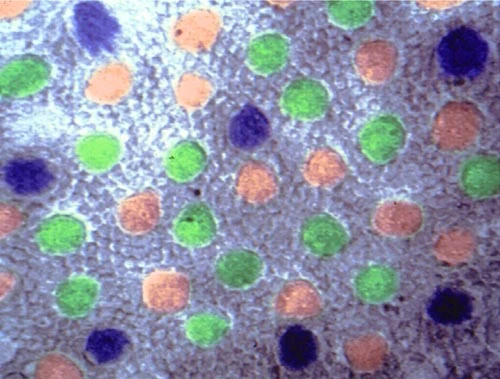
Распределение рецепторов в сетчатке бабуина . Синие колбочки были распределены регулярно в периферии сетчатки, красные и зеленые колбочки были распределены беспорядочно всюду. Плотность распределения зелёных колбочек больше, чем красных, больше чем синих.[1]
{kind=link}
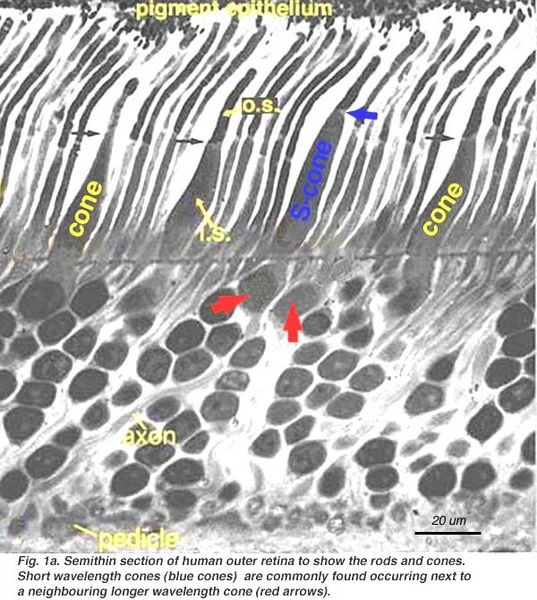
Рис.1a. Тонкий срез человеческой внешней сетчатки, где показаны палочки и колбочки (выделена «синяя» колбочка)[2]

{kind=link}
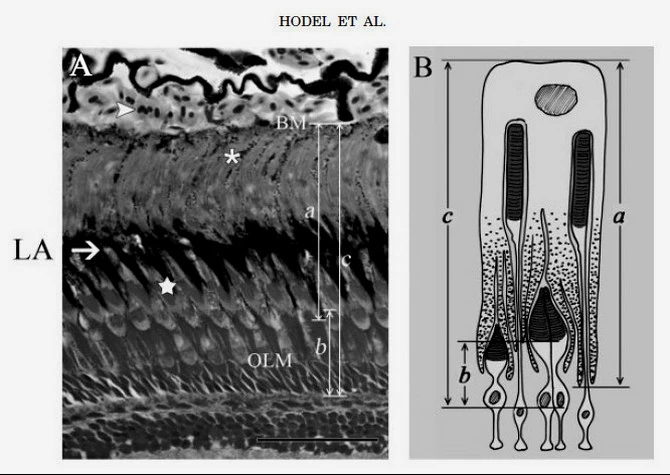
Рис. 1. Смещение гранул меланина, рецепторной части колбочек и палочек в сетчатке — при дневном освещении (дневное зрение; LA — состояние световой адаптации глаза)
Слева — микрофотография сечения сетчатки глаза рыбы, справа — схематическая прорисовка взаиморасположения органелл и фоторецепторов. Направление света, приходящего в сетчатку сквозь хрусталик: снизу-вверх.
На фото видно: днём палочки скрыты гранулами меланина; рецепторная часть колбочек выдвинута к свету (на фото — смещена вниз).[3]

{kind=link}
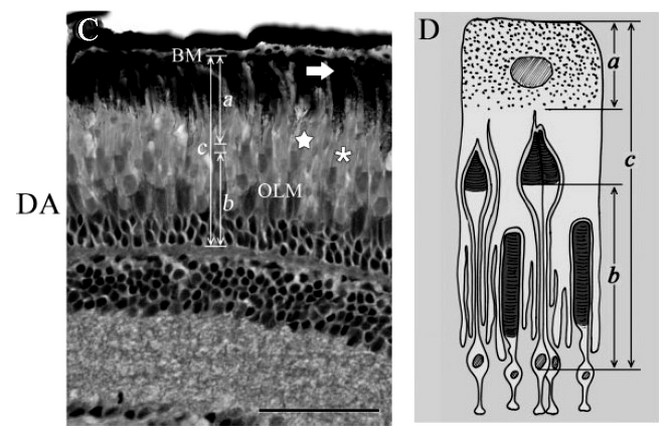
Рис. 2. Движение колбочек, палочек и гранул в условиях слабого освещения (ночное зрение; DA — темновая адаптация глаза), микрофотография сечения сетчатки глаза рыбы.
Справа — схема перемещений органелл и изменения формы фоторецепторных клеток.
На фото видно: при недостатке света гранулы меланина открыли путь света к палочкам; рецепторная часть колбочек удалена от источника света, рецепторная часть палочек — приближена к нему (на фото — смещена вниз).[4]
В вопросах цветовосприятия в последние десятилетия наметился прорыв в создании новых совершенных средств исследований, позволяющих рассмотреть живые клетки и их работу при воздействии на них видимых лучей света (цвета). Удалось рассмотреть колбочки RGB, открыть третий фоторецептор сечатки ipRGC (~2007 год), который расположен в ганглиозом слое сетчатки и связан с колбочками, палочками, мозгом. Фоторецепторы ipRGC расположены на пути перед фокальной поверхностью сетчаки, где находятся колбочки и палочки, выполняет важные фунции при цветовосприятии, в том числе в управлении работой колбочек при восприятии спектра коротковолновых лучей (синих-фиолетовых), пропуская нормальные лучи, блокируя сильнодействующие. Он содержит оптический пигмент меланопсин.
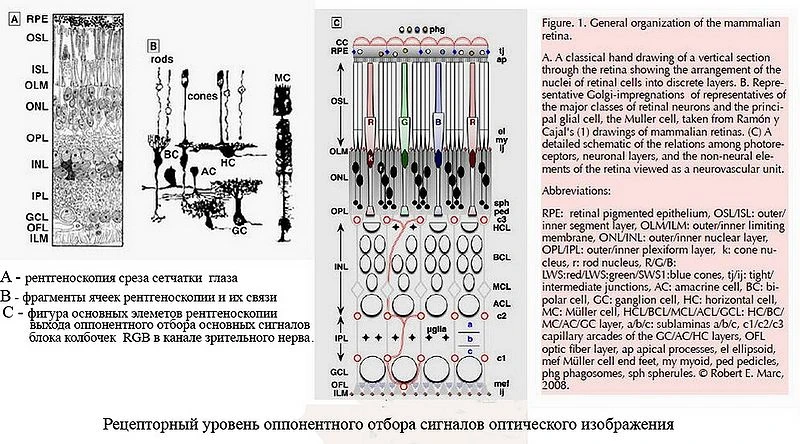
Рентгеноскопия сетчатки и зрительных отделов мозга, проведенная ещё в 1966, 2009 годах Р.Е.Марком, на атомно-молекулярном уровне подтверждает ранее сформулированые гипотезы о работе сетчатки, где заложены фоторецепторы колбочки, которые на рецепторном уровне воспринимают, трансдукцируют основные, аналоговые сигналы RGB оптического изображения (контурного) перед передачей их в высшие зрительные отделы (нейронные) головного мозга (см.Лаборатория Р.Е. Марка). Уже данная информация позволяет сформулировать основу трёхкомпонентной теории цветного зрения (некоторые участники бездоказательно пытаются её опровергнуть, ссылаясь на нелинейную теорию цветного зрения С.Ременко).
В настоящее время уже, вообще, не стоит вопрос о восприятии цвета в сетчатке глаза блоками из трёх колбочек RGB (R.Marc, 1977).[5],[6] Если говорить о восприятии синих монолучей синими колбочками, что в нелинейной теории С.Ременко, вообще, это отвергается и как альтернативой является работа палочки в блоке палочка-колбочка, то уже в семидесятых годах (1977) легко получены рентгеновские снимки синей колбочки в ряду основных колбочек зелёных и красных (см. рис.1а). (Например, доказано, что основным местом, где работают только колбочки, является желтое пятно, которое предназначено для дневного зрения).
Группа учёных Лаборатории R.Marcа уже работают над созданием атласа основных нервных клеток сетчатки и отделов головного мозга на основании электронной микроскопии их у кроликов и крыс. При этом ипользуется разработанноя мощная программа обработки данных современных микроскопов и компьютерных систем. При этом открывается возможность визуально анализировать работу здоровых и больных клеток с целью лечения целого ряда болезней (слепота, дальтонизм и др.). [7],[8].
Что важно, на рентгеноскопиии среза сетчатки (cм. Фиг.R) видно, что основные фоторецепторы колбочки и палочки в процессе формирования оптического изображения работают независимо. Так синяя колбочка при дневном зрении воспринимает диапазон синих лучей под присмотром фоторецепторов ipRGC, свзанных также со зрительными отделами головного мозга. Например, сильные сигналы ультрафиолетовых, синих лучей под действием ipRGC и мозга закрывают вообще ресницами глаза глаз от их попадания во внутрь (скорость реакции 1/2000сек) и имеются другие функции у них, связанные с трансдукцией синего сигнала.
Также последние данные исследований (см. Ретиномоторная реакция фоторецепторов) подтвердили, что палочки в дневном зрении (начиная с длин волн 498нм) вообще не участвуют. Можно возразить, что эксперимент проведен на рыбах. Но в живой природе рыбы, птицы, наземные животнве обладают зрительной системой аналогичной человеку.(Есть глаз, сетчатка, колбочки, палочки и т.д.) В результате эволюции зрителной системы глаз постоянно меняется, но суть одна и таже. Без зрительной системы многие виды не выжили бы. Наприер, у птиц (см.Цветное зрение у птиц) зрение наиболее "острое", чем у человека. Откуда, надо спросить у противников трихроматизма (сторонников нелинеёной теории зрения С.Ременко), как можно загружать Институт прикладной физики и Институт химии АН МССР, совместно с кафедрой офтальмологии Кишинёвского Государственного медицинского Университета работой напрвлениями в области цветного зрения согласно теории С.Ременко, где нелинейная модель анализа цвета позволяет представить цвет в виде чисто физических параметров на базе результатов преобразования параметров излучения на фототранзисторах, вместо живых клеток колбочек и палочек и ещё на приборах типа колориметров вместо прямого применения живых клеток, которые рассматривают на новых флюоресцентных микроскопах. Как можно извесные физические законы применить в системах живых организмов игнорируя сам объект исследования, саму живую природу зрения — живой глаз, живую сетчатку, живые клетки. Больше того, как можно такие коллетивы загружать работой заранее не перспективной, не признанной, когда вся мировая наука в области зрения, цветного зрения доказала и приняла принцип трихроматизма с учётом исследований на разных уровнях — физическом, биофизическом, биохимическом, которые не смогли обойти принцип трихроматизма - принцип цветного зрения с участием только колбочек, которые в итоге взаимно дополняли и доплняют друг другаи и в настоящее время. Как результат: в 2011 году было открыто фундаментальное, принципиальное достижение в области цветного зрения — это явление ретиномоторной реакции фоторецепторов глаза (см. рис.1, рис.2). Что после этого можно сказать? Палочки не участвуют при цветном зрении. Работают, как ранее было даказано, только колбочки. А вышеназванные институты с подачи их сотрудников, работая в системе нелинеёной теории цветного зрения, на протяжении более (2011-1975=36) 30 лет, трудились вхолостую! Не работает система «колбочка+палочка», все доказательства — подгонка результатов под нужный, не объективный резьтат. Лучше если сказать, плачевный. (Хотя отрицательный экперимент в науке также важен).
Подходы в выборе основ цветного зрения[]
| Клетки Колбочек человека | |
|---|---|

|
На основе установленных законов цветовидения за период более 200 лет было разработано несколько теорий цветового зрения. Однако, приблизительно в последние 25 лет появилась возможность непосредственно проверить эти теории методами электрофизиологии путем регистрации биоэлектрической активности одиночных фоторецепторов зрительной системы, а также расширить исследования в области взаимодействия и количества фоторецепторов сетчатки и мозга в цветовосприятии. Так в 2007 году признано открытие 1991 года эктерорецепторов ipRGC (не колбочки и не палочки, расположенных в ганглиозном слое на пути световых лучей оптического изображения в сетчатке глаза), в 2006 году был изобретен флюоресцентный микроскоп, в 2009 году учёным Р.Е.Марком повторно была проведена микроскопия среза живой сетчатки глаза человекоподобной обезъяны, где в цвете были обнаружены экстерорецепторы сетчатки, в 2011 году было открыто явление ретиномоторной реакции фоторецепторов. Уже толко на основании этого открытия можно закончить дискуссию с противниками трихроматизма. Доказано и показано на снимках, что цветное зрение воспринимается только колбочками. Палочки в цветном зрении не участвуют.
В результате все теории сошлись к одному принципу трихроматизма - цветное зрение основано на отборе трёх основных лучей света RGB колбочками в блоке не менее трёх с нормализованными спектрами чувствительностьи S,M.,L (синие. зелёные, красные).
С точки зрения нелинейной теории, начиная с 1975г. по сегодняшний день, её сторонники применяют технологию исследований на основе:
- 1)Подход чисто физический, без участия головного мозга и живых клеток колбочек, палочек и третьей недавно открытой клетки ганглиозного слоя ipRGC;
- 2)Вместо палочек и колбочек применяются фототранзисторы и исследование проводится на роботе глаза - колориметре, испльзуемый ещё в 30 годах для получения атласа красок методом тестирования сторонних наблюдателей.
- 3)Все умозаключения производят на базе данных работы фототранзисторов с разными источниками света под управлением оператора.
- 4)Все параметры цвета в нелинейной модели цветовосприятия — все получаемые световые сигналы при колориметрии не попопадают в мозг, не проходят оппонентный отбор с участием мозга и других экстерорецепторов сетчатки недавно открытых, которых не знал С.Ременко (например, см. Фоточувствительные клетки сетчатки ipRGC), и по мнению С.Ременко сигналы создаются в лаборатории и имеют строгое назначение и чёткий физический смысл.[10] [11] Как видим колориметрия цвета имеет прямой смысл для создания каталогов цветов для практических целей, например, применения в полиграфии, для производмтва промышленных красок, для художников и т.д.
{kind=link}
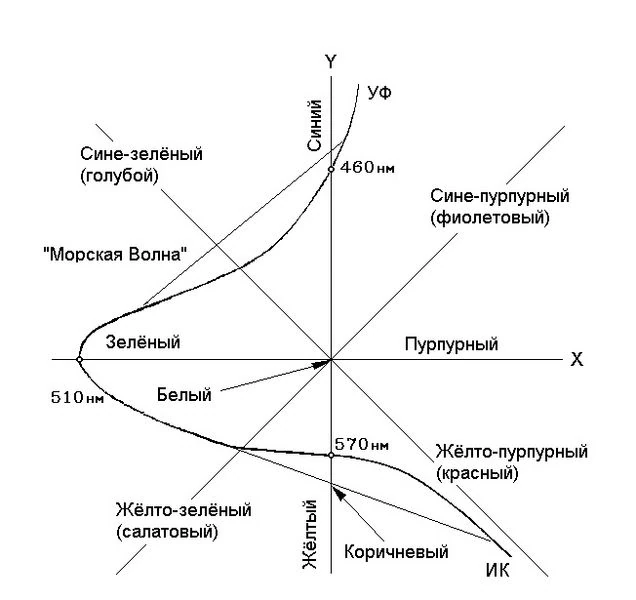
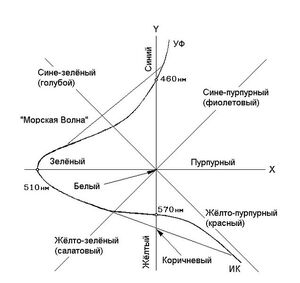
Рис. 4. Цветокоординатная система нелинейной модели и кривая видности глаза. Кривая, описывает положение спектральных цветов (непрерывного спектра полученного разложением призмой белого солнечного света) на координатной плоскости.
При этом на цветокоординатной системе нелинейной модели находятся все возможные цвета и оттенки, которые в природе и не существуют и глаз нигде их не видел (см. рис. 4). Фактически цветокоординатная система нелинейной модели представляет собой всем давно известный «цветовой круг». В центре находится белый, по периметру чёрный. Любые два противоположных (относительно центра координат) цвета равноудалённые от центра, в сумме дадут белый. Яркость — третья координата, перпендикулярная плоскости. Все существующие цвета имеют своё место в этой полусфере. На рисунке сплошной, толстой линией показано положение спектральных цветов (одинаковой яркости) на цветокоординатной системе.
Тем не менее все возможные цвета и оттенки, которые в настоящее время получают методом колориметрии, не могут сравниваться с цветом и оттенками, которые формируются в зрительных отделах головного мозга, не говоря уже о динамике восприятия. Те зрительные сигналы , которые получает мозг, вообще, несопоставимы с сигналами, которые формирует колориметр.
Цвет, полученный искусственно, оценивается среднестатистическим наблюдателем и его название вносится в каталог цветов. Боьше того, нет никакой связи с работой третьего экстерорецептора ганглиозного слоя сетчатки, не говоря о недавено открытом явлении (2011г.) как Ретиномоторная реакция фоторецепторов, которое окончательно показывает на сделанных снимках, что при цветном зрении работают только колбочки и что палочки работают только в условиях сумеречного и ночного освещения.
- Цвет — как физический параметр, оппонентно отобранный и полученный в зрительных отделах головного мозга, оценивается только индивидуально у каждого индивидуума. Спектральное же распределение всех излучений, cоздаваемое фотоприемным устройством, например, колориметром, в конечном итоге аттестуется - оценивается и сравнивается среднестатистическим на6людателем.
Взгляды сторонников трихроматизма или трёхкомпонентной теории зрения[]
Требования к теории цветного зрения[]
Факты и их объяснение
К главным фактам, раскрывающих суть трёхкомпонентной теории цветного зрения, относятся:
- Экспериментальные гистологические научные данные, проводимые при микроскопии на базе современных флюоресцентных наноскопов с использованием живых клеток (сечений сетчатки глаза) с разрешающей способностью 1-10нм, в цветовом трёхмерном пространстве на атомно-молекулярном уровне. (См. Цветное зрение у птиц);
- Колориметрия;
- Цветоведение;
- Оптика;
- Физиология и анатомия человека;
- Биохимия восприятия света;
- Генетика;
- Психология восприятия цвета.
Все указанные факторы должны быть непротиворечиво объединены с помощью наиболее понятного понятийного и математического аппарата.
Среди фактов, которые должны быть учтены:
- Данные последних исследований строения сетчатки при микроскопии (рентгеноскопии, флюоресцентной наноскопии);
- Учёт спектра чисто земных изображений, который может быть в виде двух основных компонентов:
- Цветной контраст между короткими и средними длинами длин волн спектра;
- Цветной контраст между средними и длинными частями длины волны спектра.
- Должна быть использована информация о приматах и челоеке, и чтобы извлечь и использовать эту информацию, требуется:
- Определение в видимой части соответствующей области длины волны;
- Учёт морфологии (схемы) сетчатки глаза, которая должна приспосабливаться, воспринять спектральную информацию и быть эффективно переданной в мозг.
Животные, приматы, зрительные системы которых отвечают этим требованиям, наделены способностью цветного зрения. [12],[13].
- Должно быть учтено и выдвинуто на первый план недавнее продвижение понимания трех аспектов видения цвета trichromaticу у приматов (2002г):
- 1) поведенческое значение trichromacy,
- 2) развитие и генетика фотопигментов,
- 3) относящиеся к сетчатке глаза кругообороты, которые создают красно-зеленые и синие-жёлтые цветные каналы в зрительном нерве.[14].
Принципиальная точка зрения[]

{kind=link}
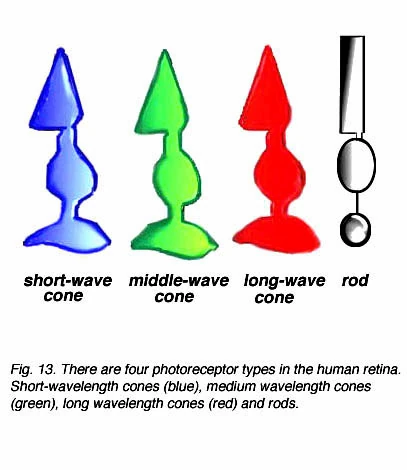
Рис. 13. Типы фоторецепторов колбочек и палочек[15]
Принцип трихроматизма или трёхкопонентный принцип цветного зрения основаны на базе физики цвета (RGB) и взаимодействия с ним живой среды обитания — биологической, а также на основе колориметрических, гистологических, генетических исследований, на основе пересмотра прежнего сложившегося процесса зрения на базе труда (2011года) учёного физика Джеральда К. Хата [16], который происходит на базе и из физики взаимодействия света с внешними долями мембран фоторецепторов сетчатки глаза, где в основном осуществляется первичное взаимодействие со светом (рецепторное). [17] [18].
Даже с точки зрения чисто физичекой (не затрагивая биофизику, биохимию и др. области) работа сетчатки укладыается в направление, которое подводит к «nanostructural», когда свет рассматривается с вопросом как электромагнитная волна в классической физике, который проходит через пространственную размерность (названной «nano-антеннами») и фильтруется в виде трёх основных цветов RGB в мембранах колбочек и палочек — образуя систему трихроматизма.


{kind=link}
Рис.Б. Восьмугольная симметрия присутствует на сетчатке глаза в 7-8° (степенях) оригинальности, где статистически плотность палочек сначала достаточна, чтобы полностью окружить каждое уменьшающееся число колбочек (нано-антена — 1колбочка, окружённая восьмью палочками).[19]
Падающий на сетчатку свет и его восприятие следует подразумевать, что при поглощении света имеет место взимоотношения в пространственной размерности элементов между смежной колбочкой и палочкой, но не в виде чистого кванта и имеется в виду, что фотоны взаимодействуют непосредственно в пределах образования — фоторецепторов: 1 колбочка:8 или 6 палочки вокруг неё!.
Последние данные Джеральда К. Хата (2010-2011) не противоречат полученным результатам известных учёных всей предыдущей более 200 летней истории цветного зрения, а только их уточняет, исправляет во взаимодействии с ним (см. Теории цветного зрения).
Фоточувствительные экстерорецепторы сетчатки[]
Фоточувствительные экстерорецепторы сетчатки — экстерорецепторы, расположенные в сетчтатке глаза, воспринимающие электромагнитное излучение в световом диапазоне длин волн. Фоторецепторы обеспечивают и создание оптического изображения — функцию зрения, и внешнюю коррекцию биоритмов сна и бодрствования, зависящую от общей освещённости.
{kind=link}
Рис.1 Структура сетчатки глаза (у приматов). (R.E.Marc, 2009, [20])
Взгляды сторонников нелинейной теории зрения[]
В 1975 году появилась нелинейная теория зрения советского ученого С. Ременко, предполагающая наличие в глазе человека только двух типов светочувствительных элементов — одного типа палочек и всего одного типа колбочек, содержащих в себе пигменты светочувствительные сразу к нескольким областям спектра (например, красных, зелёных), а также имеют нелинейность процессов формирования сигналов цветности. В отличии от всех остальных существующих на сегодня теорий она единственная, которая объясняет механизмы обработки сигналов рецепторами, поддержание баланса белого цвета и моделирует работу глаза в целом и базируется на методах исследований на базе модели глаза (колориметре). Оценка выходных сигналов проходит на уровне рецепторном, на чисто физических принципах (без учёта субъективных факторов, мозга), без учёта третъего фоторецептора ipRGC, а также ретиномоторной рекции фоторецепторов.[21] [22] [23] [24].
Однако, теория С.Ременко, базируясь на данных исследований, не связанных с живыми клетками глаза, основанная на построенной модели глаза (колориметре) без учёта работы колбочек, палочек и третьего недавно открытого фоторецептора ipRGC, не учитывающая недавно открытое явление — Ретиномоторная реакция фоторецепторов, без учёта работы мозга, где однозначно доказывается, что палочки не участвуют в цветном зрении, выдвинута модель мозаики сетчатки в виде блоков «палачка+колбочка». И в то же время, когда в 2011 году данные ретиномоторной реакции фоторецепторов на основе гистогенетических исследований на живой клетке на уровне флюоресцентной микроскопии показали, что колбочки и палочки работают изолированно друг от друга с границей раздела — освещённости в 498нм. До 498нм, в условиях сумеречного освещения работают полочки, свыше 498нм, колбочки открываются, а палочки прячутся, что также доказазал доктор М. А. Островский, Институт биохимической физики РАН им. Н. М. Эмануэля, Москва в работе [25], в настоящее время не хотят согласиться с тем, что модель исследований С.Ременко не только устарела, но тупиковая.
Применения методов генного моделирования у мышей в клетках сетчатки глаза, выделение колбочки-S, которая работает в зоне фиолетово-синего спектра с длиной волны, более 498нм, т.е. с окончаним сумеречного освещения, подтверждают теорию трихроиатизма. (Ешё в 1966 году, а такжеь в 2009 году, повторно на рентгеноскопических снимках Р.Е.Марка образцов сечения сетчатки были получены данные работы колбочек при цветном зрении.) В 2006 году получены изображения колбочек и палочек цыплёнка в цвете и в стерео, где также подтверждена многокомпонентная (четырехроматическая) теория цветного зрения. Откуда не случайно, теория С.Ременко изначально в мире науки зрительной системы не принята и очевидно из-за отсутствие в ней подходов исследований на живой клетке, серьёзно не прнималась и не рассматривалась ведущими лабораториями в мире. См. Цветное зрение у птиц, Лаборатория Р.Е.Марка, Ретиномоторная реакция фоторецепторов, Фоточувствительные клетки сетчатки ipRGC. (Для удобства читателей в качестве источников приведены статьи в Науке, где указаны первоисточники и переводы иностранных текстов на русский язык).
Данные исследований оппонирующих сторон[]
У сторонников трихроматизма[]

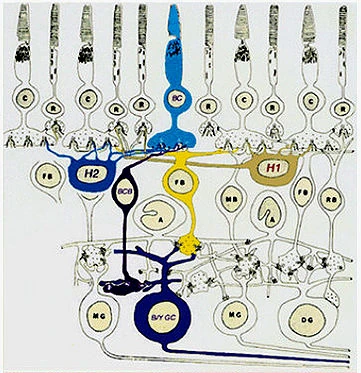
Рис. 19 Фигура «синей» колбочки сетчатки примата при электронной микроскопии и её связи с ячейками ганглиозного слоя сетчатки. [26]
В основном в области зрения основное внимание учённых уделяется вопросам исследований фоторецепторов и сетчатки, где они находятся. Главным инструментом исследований — микроскопия сетчатки. Это исследования сетчатки и её сечений с целью определения её структуры на макро- и микроуровнях, вплоть до молекулярного. Микроскопия проводится в комплексе с гистологическим и генетическим анализом структурных элементов; определеним взаимосвязей нервных клеток, фоторецепторов сетчатки, изучением мозаики блоков фоторецепторов (рентгеноская и флюоресцентная микроскопия клеток имеет разрешающую способность менее 1-10 нм). Исследование связано с появляющейся в настоящее время возможностью создания атласа связей всех типов нервных клеток сетчатки и отделов головного мозга (см. коннектомика и Human Connectome Project), на основании микроскопии приготовляемых препаратов сетчатки лабораторных животных (птиц, кроликов, голубей, черепах, крыс и др.) с применением флюоресцентной микроскопии в крайнем случае с приенением рентгеноскопии. С момента 2006 года, когда немецкие учёные создали лаборную установку флюоресцентного микроскопа, у учёных открылся доступ при исследовании живых клеток ткани в цвете с полученим стереизображений на экране мониторов микроскопа на молекулярном и атомном уровне. В связи с чем учёные увидели клетки колбочек и палочек, открыли третью клетку — фоторецептор сетчатки в ганглиозном слое ipRGC и вообще смогли поближе рассмотреть срез сетчатки в профиль. Т.е. установить связи всех клеток сетчатки в перпендикулярном сечении, т.е. напрямую посмотреть прохождение видимого отражённого или прямого луча предметных точек изображения.
Лаборатория Р.Е.Марка[]
Проведенная Р.Марком ещё в 1966 году рентгеноскопия (см. рис.1) подтверждает ранее сформулированные гипотезы о работе сетчатки, где заложены фоторецепторы колбочки, которые воспринимают, трансдукцируют основные сигналы оптического изображения перед передачей их в высшие зрительные отделы (нейронные) головного мозга. Данная информация позволяет подтвердить основные положения трёхкомпонентной теории цветного зрения (которая во многом противоречит нелинейной модели цветного зрения С. Ременко).
Открытие фоторецептора ipRGC[]
В 2007 году было обнаружено[27], что в области сетчатки, где расположены ганглионарные нейроны, залегающие в глубине сетчатки, расположен особый тип нервных клеток, ipRGC.


{kind=link}
Схема слоёв поперечного сечения сетчатки глаза. Область слоя ганглиозных клеток содержит клетки нейронной сети сетчатки, фоторецепторы ipRGC
Эти клетки функционально связанны с палочками, колбочками и мозгом, они частично обуславливают реакцию на воздействие коротковолновой части спектра (460-484 нм), непосредственно не лежащие в фокальной поверхности колбочек и палочек (см. рис.2, 4), которые рефлекторно связанны прямой и обратной связью с палочками и колбочками, и сквозь слой нервных волокон, с мозгом. Потому свет, прежде чем попасть на светочувствительные элементы, палочки и колбочки, должен пройти через слой ганглиозных нейронов ipRGC, которые одновременно являются дополнительным светофильтром, отсекающим губительное для тканей и рецепторов проникновение ультрафиолетовых лучей из УФ области спектра. Пигмент меланопсин, чувствительный к фиолетово-синему участку спектра, благодаря связям с колбочками, палочками и мозгом, через фоторецепторы ipRGC участвует в работе колбочек и палочек, и в восприятии лучей S, а также в случае прямого попадания солнечных лучей и лучей электродуговой сварки в глаз мгновенным закрыванием ресниц, а также подавлением функции восприятия фиолетово-синих лучей мембраной колбочек и палочек в оппонентном отборе сине-жёлтого участка спектра.
Рецепторный уровень фототрансдукции нервных сигналов[]


{kind=link}
Принципиальная схема организации цветного зрения (на примере сетчатки цыплёнка).
А. Фоторецептор-колбочка. Свет проходит сквозь хрусталика и до пигментов колбочки расположенных на её «дне».
Б.1-Б.4 У птиц обычно четыре типа колбочек (они «тетрахроматы»), что позволяют им синтезировать многоцветное изображение, и различать цвета лучше, чем человек. Обработка полученных разностных сигналов, происходящая в нейронной сети сетчатки (десятки типов клеток) обеспечивает возможность ясного различения тысяч цветов и оттенков.
Реакции ганглиозных клеток отражают возбуждение нескольких сотен или ещё большего числа рецепторов. Не удивительно поэтому, что встречаются ганглиозные клетки, отвечающие на раздражение любого участка большой области сетчатки. Область, раздражение которой (в любом её участке) приводит к ответной реакции данной клетки, называют рецептивным полем клетки. Изучение пространственной и временной организации рецептивных полей ганглиозных клеток показывает, что значительная модификация нервных сигналов происходит уже в сетчатке, то есть до того, как сигнал будет передан в высшие отделы мозга [28].
На основании этого вытекает, что фоторецептор ipRGC рефлекторно участвуют в ощущении восприятия света и цвета.
Известно, что глазные среды имеют разные показатели преломления и среда с большими показателями преломления световые волны с меньшей длиной волны (с более высокрй частотой электромагнитных колебаний фотонов) преломляют сильнее. Это приводит к тому, что преломляющая способность оптической системы (сила) глаза «синих» лучей с длиной волны 450 нм на 1,3Д (Диоптрии) больше, чем «красных» лучей с длиной волны 650 нм. Поэтому все лучи фокусируются на разной высоте главной оптической оси, т.е. как бы не на одной сферической фокальной поверхности сетчатки. На сетчатке лучи видимого спектра образуют резкое изображение не одно: синие, зелёные, красные лучи сфокусируются резко в трёх сферах сетчатки на разных высотах мембран сетчатки. (как бы в трёх фокальных поверхностях). Такое явление при фокусировании, зависящее от длины волны света называется хроматической аберрацией. Предполагалось, что и фоторецепторы, чувствительные к той или иной части видимого спектра, должны располагаться только на том участке сетчатки, где расположены наружные и внутренние сегменты мембран фоторецепторов — палочек и колбочек. Причём области фоторецепторов (мембрана) чувствительные к той или иной длине волны расположены на различной глубине (вдоль колбочки, в зоне мембраны = 50мкм). Однако, как показали исследования фоторецепторов сетчатки колбочек и палочек с применением флюоресцентной микроскопии живых ячеек получены новые данные, которые говорят о многокомпонентной структуре восприятия цвета рецепторами сетчатки. Объединённые в блоки колбочки образуют мозаичную структуру сетчатки, где в каждом блоке («ячейке») расположены колбочки, воспринимающие свои спектральные монолучи света, например, синий, зелёный, красный у человека (трихроматизм), у птиц (птенцов) — по четыре колбочки, (+ фиолетовый).
Также на основании опубликованных данных исследований профессора департамента офтальмологии функциональной нейроанатомии сетчатки R.E.Marcа (2009), то ещё в 1966 году при электронной, рентгеновской микроскопии сетчатки полученные данные «синих» колбочек приматов [29], что не совападает с данными С.Ременко на базе его исследований на модели глаза (колориметре), где вместо живых ячеек (колбочек и палочек) сетчатки использованы фототранзисторы и утверждается, что в сетчатке глаза в восприятии цвета участвуют блоки только из двух фоторецепторов «палочка+колбочка» (1975). Больше того, рентгеноскопия сетчатки глаза (см. фиг. ) ещё в 1966 R.E.Marcом показывает связи колбочек и палочек и фоторецепторов ipRGC, где палочки и колбочки работают независимо и просматривается трёхкомпонентая структура связей колбочек RGB и ячеек ганглиозого слоя.
По мере удаления блоков от центра жёлтого пятна меняются расстояние между фоторецепторами и сами фоторецепторы. Например, на периферии (в зоне периферийного зрения) практически в группах одни палочки.
Также в результате флюоресценции жировых капелек колбочек, где проходят процессы формирования и накопления сигналов (цветов), посылаемых в мозг, то уже на этом этапе (в сетчатке) происходит оппонентгая обработка сигналов до поступления их в зрительные отделы (на нейронном уровне) головного мозга, а также определились разновидности колбочек, подтвердающих оппонентную многокомпонентную теорию цветного зрения. Например, у птиц — «четырехроматизм» (см. Цветное зрение у птиц), у человека и некоторых приматов «трихроматизм».


{kind=link}
Рис. 10;Гистология колбочек и палочек цыпленка при флюоресцентной микроскопии
Данные иссдедования (2006 год) поставили на свои места целый ряд гипотез и предположений, противоречащих одна другой. При этом сформировано общее направление трёх и более компонентной теории цветного зрения — что существует процесс трех или более компонентного восприятия цвета на 2-х уровнях: на уровне низших отдедов зрительной системы — сетчатки совместно с клетками зрительной нервной системой головного мозга (принципы оппонентного цветового зрения), которые как позже выяснилось дополняют друг друга.[30]


{kind=link}
Рис.14a Спектры видимых лучей света и фоторецепторы[31]

{kind=link}
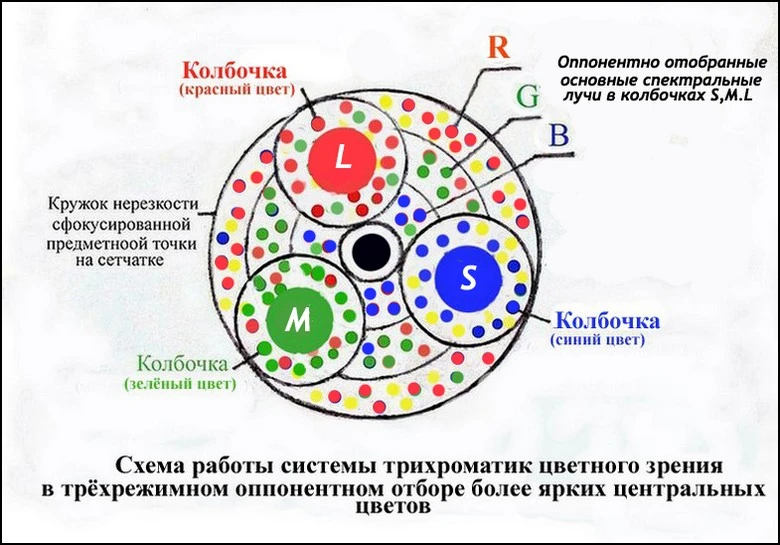
Рис.3a,Схема восприятия света и цвета в системе «трихроматик» у человека
Что касается хроматической аберрации в биологической зрительной системе глаза, то благодаря созданной природой переменной твёрдости хрусталика и роговицы, благодаря ретиномоторной реакции сетчатки и расположения фоторецепторов, например, колбочек с мембранной размерами диаметра 3-5мкм и высотой 50мкм, даёт возможность в трёхмерном пространстве сфокусироваться монохроматческим лучам света в нужном месте. При этом фокусировка каждого цвета в своей колбочке происходит в оптимальной зоне наименьшего кружка нерезкости (см. Участник:Миг/Цветное зрение, Острота зрения).
Проходящие через расположенные перед фоторецепторами капилляры лейкоциты, при просмотре в синем свете могут восприниматься как мелкие светлые движущиеся точки. Данное явление известно как энтопический феномен синего поля (или феномен Ширера)
Кроме фоторецепторных и ганглионарных нейронов в сетчатке присутствуют и биполярные нервные клетки, которые располагаясь между первыми и вторыми уровнями, осуществляют между собой контакты, а также имеющиеся горизонтальные и амакриновые клетки, которые осуществляют горизонтальные связи в сетчатке.
Между слоем ганглионарных клеток и слоем палочек и колбочек находятся два слоя сплетений нервных волокон со множеством синаптических контактов. Это наружный плексиформный (сплетеневидный) слой и внутренний плексиформный слой. В первом осуществляются контакты между палочками и колбочками посредством вертикально ориентированных биполярных клеток, во втором — сигнал переключается с биполярных на ганглионарные нейроны, а также на амакриновые клетки в вертикальном и горизонтальном направлении.
Таким образом, наружный нуклеарный слой сетчатки содержит тела фотосенсорных клеток колбочек и палочек (фокальная поверхность), внутренний нуклеарный слой содержит тела биполярных, горизонтальных и амакриновых клеток, а ганглионарный слой содержит фоточувствительные ячейки ipRGC — ганглионарные клетки, которые своими синапсами связаны с колбочками и палочками, а такжк с головным мозгом, которые при помощи обратной связи управляют колбочками и палочками. Также имеется небольшое количество перемещённых амакриновых клеток. Все слои сетчатки пронизаны радиальными глиальными клетками Мюллера.
Наружная пограничная мембрана образована из синаптических комплексов, расположенных между фоторецепторным и наружным ганглионарным слоями. Слой нервных волокон образован из аксонов ганглионарных клеток. Внутренняя пограничная мембрана образована из базальных мембран мюллеровских клеток, а также окончаний их отростков. Лишённые шванновских оболочек аксоны ганглионарных клеток, достигая внутренней границы сетчатки, поворачиваются под прямым углом и направляются к месту формирования зрительного нерва.
Каждая сетчатка у человека содержит около 6—7 млн колбочек и 110—125 млн палочек. Эти светочувствительные клетки распределены неравномерно. Центральная часть сетчатки содержит больше колбочек, периферическая содержит больше палочек. В центральной части пятна в области ямки колбочки имеют минимальные размеры (с диаметром около 2-3мкм) и мозаично упорядочены в виде компактных шестигранных блоков, но по мере центробежного удаления мозаика меняется в сторону увеличения зон блоков, в которых одноимённые колбочки, например, фиолетовые ( у птиц), строго по системе равномерного распределены и также по упорядоченой системе окружаются другими колбочками.
У сторонников нелинейной теории зрения[]
Параметры цвета в нелинейной модели цветовосприятия[]
Рис. 4. Цветокоординатная система нелинейной модели и кривая видности глаза. Кривая, описывает положение спектральных цветов (непрерывного спектра полученного разложением призмой белого солнечного света) на координатной плоскости.
Нелинейная теория цветового зрения, в отличии от трёхкомпонентной, не использует субъективных параметров цвета. Все параметры цвета в нелинейной модели цветовосприятия — все получаемые световые, видимые сигналы при колориметрии не попопадают в мозг, не проходят оппонентный отбор с участием мозга и других экстерорецепторов сетчатки. Например, недавно открытых фоторецепторов, о которых не знал С.Ременко (например, см. Фоточувствительные клетки сетчатки ipRGC). Все данные колориметрии подбираются объективно, создаются в лаборатории и имеют строгое назначение и чёткий физический смысл.[32] [33] Как видим колориметрия цвета имеет прямой смысл для создания каталогов цветов для практических целей, например, применения в полиграфии, для производмтва промышленных красок, для художников и т.д.
При этом на цветокоординатной системе нелинейной модели находятся все возможные цвета и оттенки, которые в природе и не существуют и глаз нигде их не видел (см. рис. 4). Фактически цветокоординатная система нелинейной модели представляет собой всем давно известный «цветовой круг». В центре вертикальной оси находится белый, по периметру чёрный. Любые два противоположных (относительно центра координат) цвета равноудалённые от центра, в сумме дадут белый. Яркость — третья координата, перпендикулярная плоскости. Все существующие цвета имеют своё место в этой полусфере. На рисунке сплошной, толстой линией показано положение спектральных цветов (одинаковой яркости) на цветокоординатной системе.
Тем не менее все возможные цвета и оттенки, которые в настоящее время получают методом колориметрии не могут сравниваться с цветом и оттенками, которые формируются в зрительных отделах головного мозга. Те зрительные сигналы , которые получает мозг, вообще, несопоставимы с сигналами, которые формирует колориметр. Получаемые сигналы на базе фототранзисторов-полупроводников управляются и формируются электронными системами напрямую и с любым диапазоном и в любом количестве, заложенными программами. И самое главное отличие работы зрительной системы состоит в том, что механизм формирования биосигнала в зрительной системе в процессе формирования в отличие от икусственной управляется и находится под контролем зрительных отделов головного мозга. В результате оппонентного отбора сигнала, посылаемого в мозг, обычная электронная система не в состоянии это сделать. В зрительной системе работает обратная связь двухуровневого восприятия сигнала, при котором идёт отбор наиболее чёткого, яркого луча из массы лучей, воспринимаемые уже на низшем уровне, на уровне сетчатки глаза. Например, экстерорецептор колбочка уже на этапе отбора и формировании сигнала перед выдачей его в мозг, контролируется и управляется зрительными отделами мозга. И только наше сознание участвует в оценке полученной информации, например, цвета.
Цвет же, полученный искусственно, оценивается среднестатистическим наблюдателем и его название вносится в каталог цветов. Боьше того, нет никакой связи с работой третьего экстерорецептора ганглиозного слоя сетчатки, не говоря о недавено открытом явлении как Ретиномоторная реакция фоторецепторов.
- Такм образом, цвет — с точки зрения физической, оппонентно отобранный и полученный в зрительных отделах головного мозга, оценивается только индивидуально у каждого индивидуума. Спектральное же распределение всех излучений, cоздаваемое фотоприемным устройством, например, колориметром, в конечном итоге аттестуется - оценивается и сравнивается среднестатистическим на6людателем.
Выводы[]
В итоге можно сформулировать. Существующий, общепризнанный принцип трихроматизма, трёхкомпонентной теории цветного зрения, который сформулирован на базе многовековой истории развития цветного зрения, где практически основная масса учёных приходила к одному выводу, что в основе цветного зрения со всех точек зрения основой является восприятие видимых лучей света с оппонентным отбором сигналов трёх основных лучей RGB фоторецепторами колбочками при дневном освещении - что определяет цветное зрение и в условия сумеречного и ночного освещения глаз воспринимает синие и ультрафиолетовые лучи палочками. Колбочки и палочки работают в зонах длин волн с границей в 498нм. Т.е. с меньшим значеним - работают палочки, свыше - колбочки.
Важным является то, что все исследования ученых в области трихроматизма проводились на клетках непосредственно глаз (вначале на нежживых образцах, а затем на живых клетках, например, сетчатки глаза). Откуда, все данные исследований представителями нелинейной теории зрения на роботах глаза с использованием фототранзисторов, без участия мозга не могут обобщаться, все выводы искусственные, сложные, не подтверждённые снимками микроскопии и не представляют никакого информативного интереса. В итоге нелинейная теория зрения не признана в мире. При этом в лоборатории С.Ременко не получено ни одного альтернативного снимка среза сетчатки, которые опровергли бы принцип трихроматизма. Все приведенные рисунки строятся на базе искусственных данных колориметрии и все выводы, в том числе математические не связаны с данными исследований живой клетки. Нелинеёный характер кривых нелинейной теории основан на данных колориметрии, на данных работы фототранзисторов (полупроводниковых элементов).
Так как пучок света может быть составным, состоящим из излучений многих различных длин волны, определять степень, в которой физический цвет C в Hцвет стимулирует каждую клетку колбочки, мы должны вычислить интеграл (относительно w), по интервалу [Wmin,Wmax], из C (w) *s (w), C (w) *m (w), и C (w) *l (w) (см. рис.3). Тройные из получающихся чисел связываются к каждому «физическому» цвету C (который является областью в Hцвет) к специфическому воспринятому цвету (который является единственным пунктом в R3цвет). Эта функция, как легко можно заметить, является линейной. Может также быть легко замечено, что много различных областей в «физическом» месте Hцвет может все области привести к тому же самому единственному воспринятому цвету в R3цвет. Таким образом, воспринятый цвет не единственный для одного физического цвета.
Т.е. человеческое цветное восприятие определено определенной, групповой линейной картографией от бесконечномерного места Hilbert Hцвет к 3-мерному Евклидовому месту [1]R3цвет.
См. также[]
Примечания[]
- ↑ http://prometheus.med.utah.edu/~marclab/marclab_09_outreach-vision.html
- ↑ http://webvision.med.utah.edu/photo1.html
- ↑ http://www.imls.uzh.ch/static/CMS_publications/neuhauss/literatur/pdf06/Hodel_AnatRec_06.pdf
- ↑ http://www.imls.uzh.ch/static/CMS_publications/neuhauss/literatur/pdf06/Hodel_AnatRec_06.pdf
- ↑ http://prometheus.med.utah.edu/~marclab/marclab_09_outreach-vision.html
- ↑ http://www.ncbi.nlm.nih.gov/pubmed/403607?dopt=Abstract
- ↑ http://prometheus.med.utah.edu/~marclab/marclab_09_outreach-disease.html
- ↑ http://www.amazon.com/Human-Eye-Structure-Function/dp/0878936459
- ↑ http://en.wikipedia.org/wiki/Cone_cell
- ↑ С. Ременко, «Нелинейная модель измерения цвета и уточнение терминов колориметрии», Всеакадемический семинар по проблемам стандартизации и метрологии, Ташкент, 20 — 25 ноября 1986 год, стр 41 — 42.
- ↑ С. Ременко, «Определение основных понятий в области колориметрии и измерения цветовых параметров излучения», V Всеакадемический семинар по проблемам стандартизации и метрологии Ереван, 16 — 20 ноября 1987 год, стр 58 — 59.
- ↑ ГР Jacobs, Neitz М., Deegan JF, и Neitz J. Trichromatic окрашивают видение в Новых Мировых обезьянах. Природа 382: 156†“158, 1996. [Medline]
- ↑ Neitz М. и Neitz J. Молекулярная генетика цветного видения и цветных дефектов видения. Арч Офталмол 118: 691†“700, 2000. [FreeВ FullВ Текст]
- ↑ http://physiologyonline.physiology.org/content/17/3/93.full
- ↑ http://webvision.med.utah.edu/photo2.html
- ↑ http://www.ghuth.com/
- ↑ Goldsmith, Timothy H. (July 2006). «What birds see» (PDF). Scientific American: 69-75. http://www.csulb.edu/labs/bcl/elab/avian%20vision_intro.pdf
- ↑ http://www.csulb.edu/labs/bcl/elab/avian%20vision_intro.pdf
- ↑ http://www.ghuth.com/
- ↑ http://prometheus.med.utah.edu/~marclab/Marc_Duanes_FNAR_20080815_layout.pdf
- ↑ С. Ременко, «Цвет и зрение», «Картеа Молдовеняскэ», Кишинёв, 1982 г.
- ↑ С. Ременко, «Нелинейная модель измерения цвета и уточнение терминов колориметрии», Всеакадемический семинар по проблемам стандартизации и метрологии, Ташкент, 20 — 25 ноября 1986 год, стр 41 — 42.
- ↑ С. Ременко, «Определение основных понятий в области колориметрии и измерения цветовых параметров излучения», V Всеакадемический семинар по проблемам стандартизации и метрологии Ереван, 16 — 20 ноября 1987 год, стр 58 — 59.
- ↑ С. Ременко, «Как построить глаз», журнал «Техника Молодёжи», 1981 год,№ 7, стр. 21 — 25.
- ↑ http://www.library.biophys.msu.ru/PDF/3353.pdf М. А. Островский ФОТОБИОЛОГИЧЕСКИЙ ПАРАДОКС ЗРЕНИЯ
- ↑ http://prometheus.med.utah.edu/~marclab/Marc_Duanes_FNAR_20080815_layout.pdf
- ↑ Zaidi FH, Hull JT, Peirson SN, Wulff K, Aeschbach D, Gooley JJ, Brainard GC, Gregory-Evans K, Rizzo JF 3rd, Czeisler CA, Foster RG, Moseley MJ, Lockley SW. Short-wavelength light sensitivity of circadian, pupillary, and visual awareness in humans lacking an outer retina. Curr Biol. 2007 Dec 18;17(24):2122-8. http://www.current-biology.com/content/article/abstract?uid=PIIS0960982207022737
- ↑ Ч. Пэдхем, Дж. Сондерс, «Восприятие света и цвета», Перевод с английского Р. Л. Бирновой и М. А. Островского, Издательство «Мир», Москва, 1978 год. стр. 37
- ↑ http://prometheus.med.utah.edu/~marclab/Marc_Duanes_FNAR_20080815_layout.pdf
- ↑ http://physiologyonline.physiology.org/content/17/3/93.full
- ↑ http://webvision.med.utah.edu/
- ↑ С. Ременко, «Нелинейная модель измерения цвета и уточнение терминов колориметрии», Всеакадемический семинар по проблемам стандартизации и метрологии, Ташкент, 20 — 25 ноября 1986 год, стр 41 — 42.
- ↑ С. Ременко, «Определение основных понятий в области колориметрии и измерения цветовых параметров излучения», V Всеакадемический семинар по проблемам стандартизации и метрологии Ереван, 16 — 20 ноября 1987 год, стр 58 — 59.