Пробле́ма гигантизма палеозой-мезозойской мегафауны — один из неразрешённых вопросов современной биологии, представляет из себя многократное превышение размеров и масс отдельных видов наземных животных палеозой-мезозоя по сравнению с современными человеку представителями этих видов, либо животными, занимающими аналогичную экологическую нишу[1][2][3][4].
Неоспоримые палеонтологические данные в отношении реальности гигантских насекомых карбона, титанозавров, огромных диплодоков, тероподов и птерозавров юры и мела не имеют удовлетворительного научного объяснения[5][1] вследствие скудности или противоречивости информации в отношении либо экологии упомянутых геологических периодов, либо ограниченности современных научных знаний в отношении свойств окружающей нас природы.
Проблема гигантизма[]
В 2005 году Джон Мейнард Смит, популяризировавший применение анализ размерности в биологии, в своей книге «Математические идеи в биологии» указывал на то, что в случае увеличения размеров сухопутного животного, при неизменной форме и плотности биологических тканей, масса увеличивается в пропорции . Прочность же костей зависит от площади их поперечного сечения . Таким образом, нагрузка на тело растёт быстрее, чем их прочность. А так как графики квадратного уравнения и кубической функции неизбежно пересекутся, то существует критическая точка, за которой прочности костей и мышц не хватит, чтобы удержать тело наземного животного на ногах.[6]


Необходимо помнить, что в естествознании, как в геологии, так и в биологии, уже два века господствует методологический подход, называемый «принципом актуализма», согласно которому геологические и биологические процессы, происходившие в прошлые эпохи идентичны современным, и поэтому при анализе прошлых событий постулируется, что в древности существовали те же природные закономерности, что и в современности. Этот принцип позволяет предположить, что механическая удельная прочность костей и мышц ископаемых животных принципиально не менялась на протяжении миллионов лет, и что до появления фактов, опровергающих его действенность, это допущение может использоваться в рамках рабочей гипотезы (свойство презумпции).[7]
Проверку обоснованности применения принципа актуализма в 2006 году провела группа немецких учёных во главе с Анке Кайссер-Пизалла (Anke Kaysser-Pyzalla) — материаловедом и инженером-механиком, научным руководителем Берлинского центра материалов и энергии имени Гельмгольца, профессором Рурского университета в Бохуме. В частности, были применены методы рентгеновской дифракции, протон-индуцированной рентгеновской эмиссионной (ПИРЭ) спектроскопии и некоторые другие. Сравнение первичной фиброламеллярной структуры (обеспечивающую быстрый рост) кости у зауроподов, доминирующем типе ткани в коре длинных костей, с фиброламеллярной и хаверсовой структурами костей современных крупных млекопитающих даёт основание утверждать, что кость зауроподов сохраняет свою первоначальную ориентацию кристаллитов и что её микроструктура на разных иерархических уровнях, по-видимому, такая же, как у современной кости. Таким образом, современные данные опровергают гипотезу о том, что кости динозавров зауроподов были необычайно высокопрочным материалом.[8]
Проблема успешности выполнения различных жизненных функций[]
Руководствуясь принципом актуализма проведём оценку последствий увеличения животного в 2 раза:
- все линейные размеры увеличатся вдвое (в 2 раза):

- все площади — в 2,то есть в 4 раза:


- все объёмы — в 2, то есть в 8 раз:


- а поскольку плотность остаётся неизменной, масса также вырастет в 8 раз:

- если посчитать силу мышц, то она оказывается пропорциональна не их массе, а площади их поперечного сечения:

- значит, в данном случае сила мускулатуры животного возросла в 4 раз, а его масса — в 8, то есть относительно своего веса существо стало слабее в 2 раза:

Полученная модель животного столкнётся с проблемами поддержания позы и локомоции: слишком слабые мышцы не позволят животному обеспечить успешное выполнение различных жизненных функций, таких как поиск пищи, защита от хищников и конкурирующих видов, брачные отношения, вынашивание, выхаживание и защита потомства, и т. п.[9]
Для обеспечения жизнеспособности необходимо снизить вес. И, если не уменьшать габариты тела, то это возможно лишь за счёт уменьшения массы скелета благодаря увеличению объёмов полостей в его элементах (что ведёт к нежелательному снижению предела сопротивления приложенным нагрузкам), с одновременным повышением предела сопротивления за счёт усовершенствования его геометрии.[10] В частности, по этому принципу сведена до минимума масса экзоскелета у современных насекомых и эндоскелета у современных птиц.
Вариация пропорций в совокупности с естественным отбором приводит к разнообразию животных даже в пределах одного вида и является важным условием успешности выживания таксона в целом.
Необходимо помнить, что процесс увеличения размеров тела не бесконечен.
Проблема превышения веса, соответствующего пределу прочности костей[]
Рассмотрим этот вопрос на примере современного африканского саванного слона.
Длина самца составляет примерно =6,5 м, а масса =7 тонн[11].


Допустим, что в процессе эволюции в будущем слоны увеличат свои размеры в 2 раза и достигнут длины =13 м.[12]

Напомним, что прочность костей пропорциональна площади их поперечного сечения, а площадь сечения является функцией квадрата линейного размера [13]. Поэтому при увеличении длины слона в два раза, суммарная прочность четырёх плечевых костей или четырёх бёдер (на которые распределяется вес животного) увеличится в 22=4 раза и четыре несущие кости ног будут способны обеспечивать жизненно необходимые функции 28-тонных слонов.
Увеличение реальной массы животного при сохранении его первоначальных форм пропорционально кубу линейного размера [13]. Отсюда следует, что при увеличении длины слона в 2 раза, масса слона увеличится в 23=8 раз — до 56 т.

Очевидно, что при двукратном увеличении длины слона, при условии сохранения его формы, прочность костей будет в 2 раза меньше прилагаемой нагрузки собственного тела, что естественно приведёт к разрушению элементов скелета.
Примеры вариации размеров некоторых животных[]
Двупарноногие многоножки[]

{kind=link}
Рис. 1. Артроплевра армата (реконструкция), её длина доходила до 2,5 м.
Артроплевры (лат. Arthropleura) — род ископаемых гигантских многоножек (Diplopoda), известный из отложений верхнего карбона — нижней перми (314,6—290,1 млн лет назад)[14]. Остатки обнаружены на территории Польши[15], Германии, Бельгии, Франции, Англии[14] и на северо-востоке Северной Америки[16]. Артроплевры — самые крупные из известных сухопутных беспозвоночных.[17]

{kind=link}
Рис. 2. Двупарноногая современная гигантская многоножка длиной примерно 20 см.
Тело было плоское и состояло из ряда сегментов, заходивших один за другой. У Артроплевры арматы было 30 пар ног, длина тела была 2,5 м, вес оценивается в 10 кг.[18]

{kind=link}
Рис. 3. Сравнительная размеры Меганевры (реконструкция), размах крыльев порядка 70 см и человека.
Двупарноногие многоножки, или диплоподы (лат. Diplopoda) — класс многоножек, названный так потому, что на сложных по происхождению туловищных сегментах (диплосегментах или диплосомитах) его представителей имеется по две пары ног. К 2013 году учёными описано 7842 вида, включая 5 ископаемых видов. Большинство из них — сапрофаги, питающиеся в почве растительными остатками; гораздо меньше форм питается живыми растениями. Наиболее известные представители этой группы — кивсяки.[19]
У диплопод продолговатое тело, длиной от нескольких миллиметров до 10-20 см. Оно состоит из головы, свободного шейного сегмента и сегментированного туловища.[19]
Превышение Артроплеврой линейных размеров диплопод — 12.5 раз. Различие в весе неизвестно, вследствие отсутствия этих данных в научной литературе.
Стрекозы[]


{kind=link}
Рис. 4. Крупнейшая в наши дни стрекоза Megaloprepus caerulatus, размером 19 см.
Основные статьи: Меганевры, Стрекозы
Mеганевры (лат. Meganeura) — род вымерших стрекозоподобных насекомых, живших в каменноугольном периоде (303,4-298,9 млн лет назад). Включает в себя Meganeura monyi, имевшую размах крыльев до 70 см и являющуюся наряду с Meganeuropsis permiana одним из крупнейших насекомых всех времён. Охотились они на примитивных растительноядных насекомых диктионеврид. Личинки вели наземный образ жизни и тоже были хищниками.[20][21]
Стрекозы (Odonáta) — в современности, относительно крупные насекомые, с подвижной головой, большими глазами, короткими щетинковидными усиками, удлинённым стройным брюшком и четырьмя прозрачными крыльями с густой сетью жилок. Стрекозы — активные специализированные хищники, которые питаются насекомыми, пойманными на лету. Представители отряда широко распространены по миру, встречаясь на всехматериках, исключая Антарктику.[22]


{kind=link}
Рис. 5. Кетцалькоа́тль (реконструкция) имел, по разным оценкам, размах крыльев 15 м и вес от 200 кг до 250 кг.
Размеры представителей группы современных стрекоз колеблются в довольно широких пределах. Одними из самых мелких стрекоз являются азиатские виды — Nannophya pygmaea при длине тела 15 мм и размахе крыльев 20 мм.[23] Крупнейшей современной стрекозой является Megaloprepus caerulatus, обитающая в Центральной Америке, с длиной тела до 100 мм и с размахом крыльев до 191 мм.[24]
Превышение линейных размеров Mеганевры над Megaloprepus caerulatus в 3,25 раза, а расчётная разница в весе — примерно в 34 раза.
Кетцалькоатль и современный розовый пеликан[]


.jpg){kind=link}
Рис. 6. Розовый пеликан - самая крупная водоплавающая птица. Длина тела самцов — до 185 см, а размах крыльев — до 380 см, вес — примерно 15 кг.
Основные статьи: Кетцалькоатль, Розовый пеликан
Кетцалькоа́тль (лат. Quetzalcoatlus) — один из крупнейших известных представителей семейства аждархид отряда птерозавров (Pterosauria). Единственный вид — Quetzalcoatlus northropi. Размах крыльев точно не известен из-за неполной сохранности останков, но по пропорциям птерозавров других видов (с размером до 10 м[25]) оценивается приблизительно в 12 метров (по мнению некоторых палеонтологов — до 15 м)[26], а вес — по разным оценкам, доходил до 200-250 кг[27][28]. Обнаружен в позднемеловых отложениях Северной Америки. Время обитания — верхний меловой период, примерно 70 миллионов лет назад.

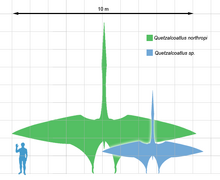
{kind=link}
Рис. 7. Сравнение размеров Кетцалькоатля northropi — зелёного цвета, Кетцалькоатля sp. — синего цвета (реконструкции) и человека.


{kind=link}
Рис. 8. Sarcosuchus (реконструкция). Его длина достигала 11-12 м, вес доходил до 8 т.


.jpg){kind=link}
Рис. 9. Гребнистый крокодил. В длину он достигает 6,7 м, а его вес доходит до 1,36 т.
Ро́зовый пелика́н (лат. Pelecanus onocrotalus) — самая крупная водоплавающая птица семейства пеликановых. Длина тела самцов до 185 см. Размах крыльев — до 380 см.[29] Вес самцов — примерно13 кг, самок — 5,1-10 кг.[30]
Превышение Кетцалькоатлялинейных размеров Розового пеликана в 4 раза, а превышение в весе — примерно в 15 раз.
Саркозух и гребнистый крокодил[]
Основные статьи: Саркозух, Гребнистый крокодил
Саркозу́х (от др.-греч. σαρκός «плоть» и σοῦχος «крокодил») — вымерший род гигантских крокодиломорфов, вопреки распространённому заблуждению, не относящийся к современному отряду крокодилов. Питался водными животными, например, рыбой, а также мелкими динозаврами. Обитал на территории современной Африки в начале мелового периода и является одним из крупнейших гигантских крокодилоподобных пресмыкающихся, которые когда-либо жили на нашей планете. Саркозух был приблизительно в 1.5-2 раза длиннее крупнейших современных крокодилов и достигал 11-12 м в длину, а масса его, как предполагают, доходила до 8 тонн.[31]
Гребни́стый крокоди́л (лат. Crocodylus porosus) — крупное современное пресмыкающееся, относящееся к семейству настоящих крокодилов отряда крокодилов. Крупнейший наземный хищник и рептилия на планете.[2]


{kind=link}
Рис. 10. Сравнение крокодиломорфов (реконструкция, Саркозух — вверху, фиолетового цвета, современного гребнистого крокодила — нижнее изображение салатового цвета) и человека.


{kind=link}
Рис.11. Гиганотозавр (реконструкция). По обнаруженным фоссилиям, длина крупнейшей особи оценивается в 13,2 м, а вес — в 13,8 т.


{kind=link}
Рис. 12. Белый медведь. Длина доходит до 2,6 м, а вес — до 800 кг.
Наиболее крупный представитель своего отряда, а также самый большой наземный или прибрежный хищник. Самцы этого вида могут достигать длины семи метров при массе примерно 1360 килограмм;[32] единственные современные крокодилы, регулярно вырастающие до 6,7 м в длину.[33]
Превышение линейных размеров Sarcosuchus над Гребнистым крокодилом — примерно в 2 раза, а превышение в весе — примерно в 8-10 раз.
Гиганотозавр и современный белый медведь[]


{kind=link}
Рис. 13. Сравнение Гиганотозавра (реконструкция) белого медведя и человека.
Основные статьи: Гиганотозавр, Белый медведь
Гаганотоза́вр— крупный плотоядный динозавр из верхнемеловой эпохи из семейства кархародонтозаврид надсемейства аллозавроидов. Описан аргентинским палеонтологом Родольфо Кориа в 1995 году по почти полному скелету сеноманского яруса (99,6-93,5 млн лет назад[2]]) из формации Канделерос (Рио Лимай) вблизи от провинции Неукен (Аргентина).
По длине тела (голотип имеет 12,2—12,5 м[3][4], крупнейший известный экземпляр доходит до 13,2 м[5]) был самым крупным представителем теропод, наравне с тираннозавром (крупнейший полный скелет которого составляет 12,3-13,2 м, крупнейшие фрагментарные окаменелости превышают 14 м) и спинозавром (крупнейшая особь, чей размер возможно адекватно оценить, имеет около 15 м в длину)[6.
Весил гиганотозавр около 7-8 тонн[note-7|[7]], хотя существуют оценки, указывающие массу крупнейшего известного образца гиганотозавра в целых 13,8 тонн[8].
В качестве условного сравнения мы можем выбрать белого медведя, поскольку он занимает в современности аналогичную гиганотозавру экологическую нишу крупнейших сухопутных млекопитающих хищников.


{kind=link}
Рис. 14. Патаготитан (реконструкция скелета). Длина базального молодого взрослого самца оценивается в 37 м с приблизительным весом 69—76 тонн.
Белый медведь — хищное млекопитающее семейства медвежьих, близкий родственник бурого медведя. Самцы белых медведей достигают 2,4-2,6 м от носа до хвоста и обычно весят до 600 кг, но могут достигать 800 кг. Самки имеют размеры около 2 м и, как правило, составляют около половины веса самцов.[34]
Превышение линейных размеров линейных размеров Гиганотозавра над линейными размерами белых медведей — примерно в 4-5 раз, а в весе — примерно в 10 раз.
Патаготитан и современные слоны[]


{kind=link}
Рис. 15. Африка́нский сава́нный слон — наиболее крупное наземное млекопитающее. Длина тела достигает 6-7,5 м, высота в плечах (наивысшая точка тела) — 3-3,8 м. Средняя масса тела у самок 5 тонн, самцов — 7 тонн.


{kind=link}
Рис. 16. Сравнение размеров Патаготитана (реконструкция), африканского саванного слона и человека.
Основные статьи: Патаготитан, Саванный слон
Патаготита́н — это род титанозавровых зауроподов из формации Серро Барцино в провинции Чубут, Патагония, Аргентина. Род содержит один вид, известный от нескольких особей: Patagotitan mayorum, впервые анонсированный в 2014 году, а затем достоверно названный в 2017 году Хосе Карбаллидо, Диего Полом и его коллегами. Современные исследования оценили длину типового образца молодого взрослого самца в 37 м и с приблизительным весом 69—76 тонн.[35]
Мы можем условно выбрать слоновых для сравнения, поскольку они занимают в современности аналогичную Патаготитану экологическую нишу. При этом необходимо помнить, что крупные млекопитающие наших дней спят стоя, поскольку, для того чтобы подняться с земли, им необходимо приложить все имеющие силы. А самое крупное современное сухопутное животное слон — не способен прыгать.[36]
Африка́нские сава́нные слоны́ — наиболее крупные наземные млекопитающие. Длина тела достигает 6-7,5 м, высота в плечах (наивысшая точка тела) — 3-3,8 м. Средняя масса тела у самок 5 тонн, самцов — 7 тонн.[37]
Азиа́тские (или инди́йские) слоны́ по размерам уступают африканским саванным слонам, однако их размеры тоже внушительны — старые особи (самцы) достигают массы 5,4 тонны при росте 2,5-3,5 м, длине тела — 5,5-6,4 м, хвоста — 1,2-1,5 м. Самки мельче самцов, весят в среднем 2,7 тонны. Самым мелким является подвид с Калимантана (масса порядка 2 т).[37]
Самцы самого низкорослого вида, лесного слона, достигают высоты плеча 2,-3,0 м. Самки меньше, около 1,8-2,4 м в плечах. Их вес доходит до 2-4 тонн.[37]
Превышение линейных размеров Патаготита́на над линейными размерами африканских сава́нных слонов — примерно, в 5-6 раз, а разница в весе достигает 10-11 раз.
Попытки решения проблемы[]
Отрицание проблемы гигантизма палеозой-мезозойской мегафауны[]
Статью 1985 года физика Хокканена «Размер самого крупного наземного животного» (Hokkanen J. E. I. «The Size of the Largest Land Animal»)[38] некоторые палеонтологи учитывают[12] как теоретическое обоснование точки зрения об отсутствии проблемы превышения веса представителей мезозойской мегафауны, соответствующего пределу прочности их костей. В частности, расчёты Хоканен показывают, что предел прочности костей современных животных допускает возможность существования способных двигаться сухопутных существ в диапазоне масс от 100 т до 1000 т. При этом возможность ещё большей массы, в случае новых адаптаций, не исключается.[38]
Вместе с тем, объективные данные содержат сведения о самом крупном из когда либо зарегистрированном сухопутном животном наших дней — африканском саванном слоне, расчётный вес которого был определён в 12,24 тонн (животное было застрелено 7 ноября 1974 года и описано в Анголе; этот гигант имел 4,16 метров в длину и 3,96 метров в высоту)[39].
Хоканен в своей статье оставляет без ответа заданный себе вопрос: "Почему же тогда не было таких гигантских животных, которые были предсказаны [произведенной им самим] экстраполяцией?"[38]
Гипотеза повышенного содержания кислорода в воздухе[]
Гипотеза повышенной концентрации кислорода в воздухе, как причина гигантизма насекомых верхнего карбона — нижней перми (314,6-290,1 млн лет назад), кроме того, хорошо объясняет ускоренное образование каменного угля на поверхности нашей планеты, а также переход части земноводных на сушу, что привело к появлению первых рептилий[40].
Дыхательная система большинства насекомых представлена множеством трахей, пронизывающих всё тело и открывающихся наружу с помощью дыхалец (стигм), которые регулируют поступление воздуха. Концевые ветви трахей заканчиваются звёздчатой трахейной клеткой, от которой отходят тончайшие трахейные капилляры (трахеолы). Трахейная система насекомых может быть открытой (типична для многих наземных свободно живущих видов или для водных насекомых, дышащих атмосферным кислородом) либо замкнутой. При втором типе трахеи имеются, но дыхальца отсутствуют. Этот тип характерен для водных или эндопаразитических насекомых, дышащих кислородом растворённом в воде или в полостной жидкости хозяина.[41]
Например, большую часть жизни личинки стрекоз не могут дышать атмосферным воздухом и для дыхания используются жабры расположенные на конце тела. Жабры могут быть внутренними (ректальными — в прямой кишке ) или наружными (каудальными). На брюшке личинок большинства равнокрылых стрекоз (Zygoptera) развиты хвостовые трахейные жабры, которые могут иметь вид листовидные прозрачных пластинок или овальных капсуловидных (пузыревидных) придатков. У личинок разнокрылых стрекоз (Anisoptera) трахейные жабры расположены в специальной камере в прямой кишке (ректальные жабры), которая периодически заполняется водой.[42]
Логическим продолжением гипотезы о повышенном содержании кислорода в воздухе в верхнем карбоне является еще одна гипотеза, которая связывает исчезновение насекомых-гигантов в результате падения процентного содержания кислорода в воздухе в нижней перми (массовое пермское вымирание, примерно 252 млн лет назад)[40]. Расцвет мира динозавров начался после массового пермского вымирания. Они доминировали на Земле в мезозойскую эру — в течение более 160 миллионов лет, начиная с позднего триасового периода (приблизительно 225 млн лет назад[43]) до конца мелового периода (66 млн лет назад). Исследования показывают, что уровень кислорода в этот период был ниже современного — примерно 15%[44].
Исходя из палеонтологических данных, изменение газового состава атмосферы Земли оказало влияние только на гигантизм насекомых.
Теория вариации гравитации в геологическом прошлом[]
Теория вариации гравитации в геологическом прошлом нашей планеты разрабатывалась начиная с начала текущего тысячилетия различными специалистами независимо друг от друга.
James Maxlow — признанный эксперт-геолог, в своей монографии "Terra Non Firma Earth" предоставил ряд доказательств в пользу гипотезы растущей Земли. В том числе, он утверждал, что гравитация в палеозой-мезозое была в два раза ниже современного её значения. Указанное обстоятельство, по его мнению и является причиной невиданного гигантизма мегафауны той эпохи. А последующее их вымирание — является вследствие роста нашей планеты и, как следствие, увеличения силы гравитации.[45]
Другой независимый исследователь, Stephen W. Hurrell — инженер-конструктор, написал несколько резонансных статей по проблеме гигантизма мезозойской мегафауны. В 2011 году вышло 3-е издание его книги "Dinosaurs and the Expanding Earth". Stephen W. Hurrell — горячий сторонник гипотезы гравитации, как причины гигантизма животных мезозоя.[46]
В 2015 году российские учёные, биолог А. Л. Дроздов и физик-теоретик А. С. Бурундуков опубликовали две статьи "Гигантские ящеры — палеонтологический вызов междисциплинарному синтезу" и "Палеонтологический парадокс в контексте глобального эволюционизма", в которых они представили математическую модель и доказали правильность гипотезы вариации гравитации в геологическом прошлом нашей планеты, переведя эту гипотезу в разряд теорий[47][48][49]
Вместе с тем, необходимо признать, что все попытки объяснения более слабой гравитации в геологическом прошлом нашей планеты входят в противоречие с геотектоникой, палеогеографией, геофизикой и др[47][50].
Междисциплинарное противоречие между палеонтологическим данными в отношении в 2 раза более слабой, по сравнению с современным её значением, гравитации в геологические периоды с карбона по мел, и содержанием других разделов геологии, а также астрофизики, которые отрицают возможность осуществления такого рода вариации гравитационного поля получило название палеонтологический парадокс[51].
См. также[]
- Зауроподы
- Размеры динозавров
- Биомеханика динозавров
- Физиология динозавров
- Палеонтологический парадокс
- Мел-палеогеновое вымирание
- Гипотеза расширяющейся Земли
- Гипотеза изначально гидридной Земли
- Парадокс слабого молодого Солнца
Литература[]
- Нэйш Д., Барретт П. Динозавры: 150 000 000 лет господства на Земле / Переводчик Рыбаков К. Научный редактор Аверьянов А., д-р биол. наук. — 1. — ООО «Альпина нон-фикшн», 2018. — 223 с. — [книг/9785001390107|ISBN 978-5-0013-9010-7]
- Fredric Heeren. Dinosaurs: Rise of the titans (англ.) // Nature. — 2011-7. — Vol. 475, iss. 7355. — P. 159—161. — [стандартный серийный номер|ISSN] 1476-4687 0028-0836, 1476-4687. — .
- Dinosaur Research: Chew and Stay Small — Universität Bonn (англ.). www.uni-bonn.de. Дата обращения 8 июля 2019.
- Yates A. M., Wedel M. J., Bonnan M. F. The early evolution of postcranial skeletal pneumaticity in sauropodomorph dinosaurs // Acta Palaeontologica Polonica. — 2012. — № 57 (1). — С. 85-100. — [[2]] 1732—2421
- Berner R. A. The carbon and sulfur cycles and atmospheric oxygen from middle Permian to middle Triassic // Geochimica et Cosmochimica Acta. — 2005-07-01. — Т. 69, вып. 13. — С. 3211-3217. — [[3]]0016-7037. — .
- Yates A. M., Bonnan M. F., Neveling J., Chinsamy A., Blackbeard M. G. A new transitional sauropodomorph dinosaur from the Early Jurassic of South Africa and the evolution of sauropod feeding and quadrupedalism // Proceedings of the Royal Society B: Biological Sciences. — 2010-03-07. — Т. 277, вып. 1682. — С. 787—794. — .
- Cerda I. A., Garrido A., Pol D. A New Sauropodomorph Dinosaur from the Early Jurassic of Patagonia and the Origin and Evolution of the Sauropod-type Sacrum (англ.) // PLOS ONE. — 2011-01-26. — Vol. 6, iss. 1. — P. e14572. — [[4]] 1932-6203. — .
- Paul, Gregory S. "Dinosaur models: the good, the bad, and using them to estimate the mass of dinosaurs." DinoFest International Proceedings. Philadelphia: Academy of Natural Science Philadelphia, 1997.
- https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4714876/
Ссылки[]
- Yates A. Necks question ... how did the biggest dinosaurs get so big? (англ.). The Conversation. Дата обращения 28 июля 2019.
- Was weaker gravity responsible for large dinosaur size? (англ.). Galileo’s Pendulum (25 February 2013). Дата обращения 9 июня 2019.
Примечания[]
- ↑ 1,0 1,1 Clemente C. J. , Dick T. J. M. Where Have All the Giants Gone? How Animals Deal with the Problem of Size // PLOS Biology. — 2017-11-01. — Т. 15. — С. e2000473.
- ↑ Sander P. M. , Witzel U., Preuschoft H., Läbe S., Barciaga T. Quantitative Interpretation of Tracks for Determination of Body Mass // PLOS ONE. — 2013-10-30. — Т. 8. — С. e77606.
- ↑ Jürgen Hummel, Daryl Codron, Dennis W. H. Müller, Patrick Steuer, Marcus Clauss Herbivory and Body Size: Allometries of Diet Quality and Gastrointestinal Physiology, and Implications for Herbivore Ecology and Dinosaur Gigantism // PLOS ONE. — 2013-10-30. — Т. 8. — С. e68714.
- ↑ Sander P. M. An Evolutionary Cascade Model for Sauropod Dinosaur Gigantism - Overview, Update and Tests // PLOS ONE. — 2013-10-30. — Т. 8. — С. e78573.
- ↑ Jennifer Welsh | June 4, 2012 03:00pm ET. Giant Insects Shrunk As Birds Entered Prehistoric Skies. Live Science. Проверено 28 июля 2019.
- ↑ Смит Дж. Математические идеи в биологии / Перевод с английского А. Д. Базыкина По редакцией и с предисловием канд. физ.-мат. наук Ю. И. Гильдермана. — 1. — Москва: МИР, 1970. — С. 24. — 179 с. — ISBN УД 681, 142 : 57 + 61.
- ↑ Menzel C. Actualism. Metaphysics Research Lab, Stanford University (17 июня 2000).
- ↑ Pyzalla A. R., Sander P. M., Hansen A.,. Ferreyro R, Yi S.-B. Texture analyses of Sauropod dinosaur bones from Tendaguru // Materials Science and Engineering: A. — 2006-11-10. — Т. 437. — С. 2–9.
- ↑ Hokkanen J. E. I. The size of the largest land animal // Journal of Theoretical Biology. — 1986-02-21. — Т. 118. — С. 491–499.
- ↑ Sander P. M. et al. Biology of the sauropod dinosaurs: the evolution of gigantism // Biological Reviews of the Cambridge Philosophical Society. — 2011. — № 86(1). — С. 117–155.
- ↑ Nowak R. M. Walker’s Mammals of the World. — 6. — Baltimore: The Johns Hopkins Univ. Press, 1999. — С. en. — 2015 с. — ISBN 9780801857898. (см. ISBN )
- ↑ 12,0 12,1 Carballido J. L. et al. A new giant titanosaur sheds light on body mass evolution among sauropod dinosaurs // Proceedings of the Royal Society B: Biological Sciences. — 2017. — № 284(1860). — С. 20171219.
- ↑ 13,0 13,1 Мэйнард Смит Дж. Математические идеи в биологии / Перевод с английского Базыкина А. Д.. Под редакцией и с предисловием канд. физ.-мат. наук Гильдермана Ю. И.. — 1. — Москва: Мир, 1970. — С. 15—20. — 179 с. — ISBN УД 681, 142 : 57 + 61. (см. ISBN )
- ↑ 14,0 14,1 Fossilworks: Arthropleura. fossilworks.org. Проверено 7 июля 2019.
- ↑ Pacyna G., Florjan S., Borzêcki R. New morphological features of Arthropleura sp. (Myriapoda, Diplopoda) based on new specimen from the upper Carboniferous of Lower Silesia (Poland) // Annales Societatis Geologorum Poloniae. — 2012. — № 82. — С. 121—126.
- ↑ The Excitement of Discovery (4 февраля 2012). Проверено 7 июля 2019.
- ↑ Lothar Schöllmann Der größte Gliederfüßer der Erdgeschichte – Reste von Arthropleura aus Ibbenbüren // Archäologie in Westfalen-Lippe. — 2013. — С. 19–21.
- ↑ McGhee Jr. G. R. When the Invazion of Land Failed. The Legacy of the Devonian Extinctions. — 1. — New York: Columbia University Press, 2013. — С. 238. — 318 с. — ISBN 978-0-231-16056-8. (см. ISBN )
- ↑ 19,0 19,1 Zhi-Qiang Zhang Phylum Arthropoda. In : Zhang, Z.-Q. (Ed.) Animal Biodiversity: An Outline of Higher-level Classification and Survey of Taxonomic Richness (Addenda 2013) // Zootaxa. — 2013-08-30. — Т. 3703. — С. 17–26.
- ↑ Fossilworks: Meganeura. fossilworks.org. Проверено 7 июля 2019.
- ↑ Rake Matthew. Prehistoric Ancestors of Modern Animals. — 1. — Hungry Tomato, 2017. — 20-22 с. — ISBN 978-1512436099. (см. ISBN )
- ↑ Suhling F. et al. Order Odonata / In Thorp, James; Rogers, D. Christopher. — 4. — Academic Press, 2015. — С. 893–927. — ISBN 9780123850263. (см. ISBN )
- ↑ Wilson K. D. P. Hong Kong Dragonflies. — 1. — Hong Kong: Urban Council of Hong Kong, 1995. — С. 155—157. (см. ISBN )
- ↑ Wilson K. Dragonfly Giants // Agrion // The Worldwide Dragonfly Association. — 2009. — № 13 (1). — С. 29—31.
- ↑ https://www.nature.com/articles/ncomms4567?
- ↑ Langston W. Pterosaurs. — 1. — Scientific American, 1981. — С. 122—136. — 244 с. (см. ISBN )
- ↑ Paul G. S., (2002). Dinosaurs of the Air: The Evolution and Loss of Flight in Dinosaurs and Birds. Johns Hopkins University Press. p. 472. ISBN .
- ↑ Witton M. P.; Habib M. B.; Laudet V. (15 November 2010). "On the Size and Flight Diversity of Giant Pterosaurs, the Use of Birds as Pterosaur Analogues and Comments on Pterosaur Flightlessness". PLoS ONE. 5 (11): e13982. doi:10.1371/journal.pone.0013982.
- ↑ del Hoyo J., Elliott A., Sargatal J., Christie D. A., de Juana E. Great White Pelican (Pelecanus onocrotalus) (англ.). Lynx Edicions. Проверено 7 июля 2019.
- ↑ Nelson J. B. Pelicans, Cormorants, and their Relatives. — 1. — Oxford, UK: Oxford University Press, 2006. — 680 с. — (Bird Families of the World). — ISBN 9780198577270. (см. ISBN )
- ↑ Boubé Gado, Christian A. Sidor, Hans C. E. Larsson, Paul C. Sereno The Giant Crocodyliform Sarcosuchus from the Cretaceous of Africa // Science. — 2001-11-16. — Т. 294. — С. 1516–1519.
- ↑ Michael Bright. The Frog with Self-Cleaning Feet: . . . And Other Extraordinary Tales from the Animal World. — 2. — Skyhorse, 2013. — 304 с. — ISBN 978-1626360754. (см. ISBN )
- ↑ Grahame Webb, S. Charlie Manolis. Crocodiles of Australia. — Sydney, N.S.W. ; London : Reed New Holland, 1998. — ISBN 9781876334260. (см. ISBN )
- ↑ Polar Bear. U.S. Fish & Wildlife Service.
- ↑ Carballido J. L. et al. A new giant titanosaur sheds light on body mass evolution among sauropod dinosaurs // Proceedings of the Royal Society B: Biological Sciences. — 2017. — № 284(1860). — С. 20171219.
- ↑ Умеют ли слоны бегать и прыгать? (рус.). www.vokrugsveta.ru. Проверено 7 июля 2019.
- ↑ 37,0 37,1 37,2 Nowak R. M. Walker’s Mammals of the World. — 6. — Baltimore: The Johns Hopkins Univ. Press, 1999. — С. en. — 2015 с. — ISBN 9780801857898. (см. ISBN )
- ↑ 38,0 38,1 38,2 Hokkanen J. E. I. The size of the largest land animal // Journal of Theoretical Biology. — 1986-02-21. — Т. 118, вып. 4. — С. 491–499. — [стандартный серийный номер|ISSN] 0022-5193. — [[1]]:10.1016/S0022-5193(86)80167-9.
- ↑ Largest mamal on land. Guinness World Records.
- ↑ 40,0 40,1 Lockley M., Meyer C. Dinosaur Tracks and Other Fossil Footprints of Europe. — 2. — New York: Columbia University Press, 2000. — С. 32. — 323 с. — [0-231-10710-2].
- ↑ Тыщенко В. П. Физиология насекомых. — 1. — Москва: Высшая школа, 1986. — С. 33—57. — 303 с. (см. ISBN )
- ↑ Скворцов В. Э. Стрекозы Восточной Европы и Кавказа: Атлас-определитель. — 1. — Москва: Товарищество научных изданий КМК, 2010. — С. 27—70. — 624 с. — ISBN 978-5-87317-657-1. (см. ISBN )
- ↑ Oldest evidence of dinosaurs found in Polish footprints (англ.). American Museum of Natural History. Проверено 22 июля 2019.
- ↑ Tappert R., McKellar R. C., Wolfe A. P., Tappert M. C., Ortega-Blanco J. Stable carbon isotopes of C3 plant resins and ambers record changes in atmospheric oxygen since the Triassic // Geochimica et Cosmochimica Acta. — 2013-11-15. — Т. 121. — С. 240–262.
- ↑ James Maxlow. Terra Non Firma Earth. - Oneoff Publishing.com, 2005-01-07. - 255 p. - ISBN 9780952260325.
- ↑ Hurrell S. W. Dinosaurs and the Expanding Earth. - 3. - South Wirral, United Kingdom: One-off Publishing. 2011. - 218 c. - ISBN 0952260379. - ISBN 9780952260370.
- ↑ 47,0 47,1 Бурундуков А. С., Дроздов А. Л. Гигантские ящеры – палеонтологический вызов междисциплинарному синтезу // Биота и среда заповедников Дальнего востока. — 2015. — № 5. — С. 93-112. — ISSN 2227-149X
- ↑ Бурундуков А. С., Дроздов А. Л. Палеонтологический парадокс в контексте глобального эволюционизма (рус.) // Программа и материалы научной конференции, посвященной памяти академика В.Л. Касьянова. — 6-7 октября 2015 г. — С. 4-9.
- ↑ Эксперты: гипотеза о вымирании динозавров из-за роста земного притяжения несостоятельна. ТАСС.Дата обращения 11 июня 2019.
- ↑ Эксперты: гипотеза о вымирании динозавров из-за роста земного притяжения несостоятельна. ТАСС.Дата обращения 11 июня 2019.
- ↑ Бурундуков А. С., Дроздов А. Л. Палеонтологический парадокс в контексте глобального эволюционизма (рус.) // Программа и материалы научной конференции, посвященной памяти академика В.Л. Касьянова. — 6-7 октября 2015 г. — С. 4-9.