Эволюция цветного зрения рассматривается, как часть естественного процесса биологической эволюции, связанного с естественным отбором, мутациями и селекцией некоторых наиболее подходящих для выживания особей и видов. Хорошее зрение даёт видам определённые преимущества в условиях обитания на поверхности земли, в верхних слоях океана и морей, в воздухе (птицы и насекомые). В то же время виды, обитающие под землёй, в глубине океанов утратили органы зрения, компенсировав их редукцию развитием других органов чувств.
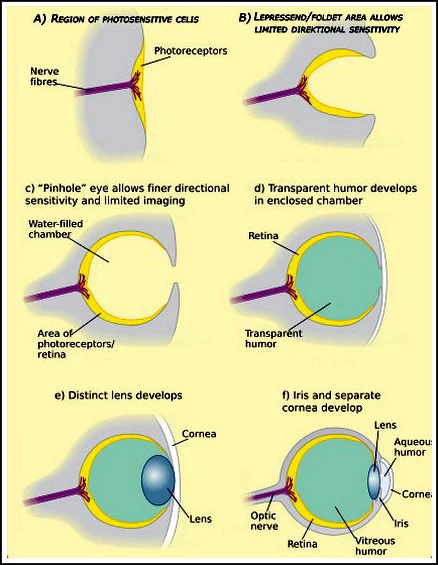
Развитие устройства зрительного аппарата[]
{kind=link}
Эволюция зрительной системы
Видимый диапазон электромагнитных волн занимает очень узкий участок всего электромагнитных волн, между ультрафиолетовым и инфракрасным диапазоном. «Видимость» — понятие чисто человеческое, такие лучи человек и многие представители мира животных воспринимают (видят) с помощью глаз. Кроме человека, приматов зрительной системой обладают многие обитатели Земли, которые ориентируются в пространстве прежде всего с помощью зрения; большинство из них в результате эволюции зрительной системы и всего природного мира так или иначе воспринимают цвета.
Свето- и цвето-чувствительность[]
Цвет — один из множества световых и других раздражителей окружающей среды, воспринимаемых зрительной системой. Видимый участок спектра обеспечивает всё разнообразие цветов и оттенков, и позволяет животным эффективно ориентироваться в окружающем мире. Несмотря на то, что он очень узок — видимая область занимает участок всего лишь от 380 нм (или 0,38 миллионной доли метра) до 780 нанометров, природа же использует его максимально полно. Для живых существ самым информативным участком спектра является видимый.
Ультрафиолетовые лучи (УФ) частично доходят до земли и одинаково сильно поглощаются как живой, так и неживой средой, материальным миром. Эти лучи человек не видит, в силу того, что они с длиной волны меньше границы 498нм, когда палочки находятся не в зоне цветного видения, а в зоне сумеречного и ночного видения. Откуда, если бы человек видел только их, то все предметы выглядели бы серо-чёрными. Инфракрасные лучи (ИК) для жизнедеятельности организма также малоинформативны. Они частично поглощаются атмосферой, при этом спектральный состав дошедших до земли ИК лучей сильно зависит от её состояния и непрерывно меняется. Если бы организм воспринимал также эту область спектра, то нестабильность погоды привела бы к тому, что цвет предметов беспрерывно менялся бы в зоне инфракрасного излучения и все окружающие предметы светились бы разными оттенками ИК лучей, что вряд ли организм сумел бы выдержать такой хаос предметов, светящихся красноватыми оттенками (диапазон волн между красным концом видимого света (с длиной волны λ = 0,74 мкм) и микроволновым излучением (λ ~ 1—2 мм). Инфракрасные лучи позволили бы живым существам различать, например, различные виды белков и т.д.
Видимая область спектра для организмов наиболее информативна. Это информация, например, о том, спелый плод или нет, можно ли его съесть или следует воздержаться. То есть это большая часть необходимой живому организму информации. Света достаточно, солнце излучает его достаточно, атмосфера почти не поглощает свет, при этом его спектральный состав в течение дня почти не меняется. Красный плод остается красным весь день и даже на следующий. Свет - это привязка к источнику жизни, к Солнцу.
Для человека видимый диапазон выглядит, как разукрашенный всеми цветами радуги.
Некоторые эволюционные вехи[]
Зрение насекомых[]
Зрение позвоночных[]
Цветовое зрение у птиц[]
Зрение у млекопитающих[]
Считается, что предки современных млекопитающих перешли к ночному образу жизни, и утратили большую часть способности к цветоразличению, сохранив лишь 2 типа колбочек и один тип палочек.
Зрение у приматов[]
Современные гипотезы эволюции цветного зрения рассматривают цветное зрение приматов, как процесс неогенеза, когда редуцированное, дихроматическое, т.е. уменьшившее свои возможности по сравнению со зрительными возможностями их предков, зрение приматов развилось вновь, но только по несколько иному эволюционному плану. Считается, что предки современных приматов перешли к дневному образу жизни на деревьях, что сопровождалось потребностью в различении спелых и неспелых плодов, оттенков зелёного цвета листвы и т.д.
Зрение у динозавров[]
Недавно вышла статья, посвящённая исследованию возможных вариантов цветовосприятия у динозавров.
Причины формирования узкого диапазона "видимого" света[]
Приходящие из космоса рентгеновские лучи задерживаются атмосферой. Организму они ни о чем сказать не могут.
Ультрафиолетовые (УФ) лучи частично доходят до земли. Эта часть спекра сильно рассеивается атмосферой, при этом они одинаково сильно поглощаются как живой, так и неживой материей. Для человека они принесли бы крайне мало полезной информации. В УФ части спектра все предметы выглядели бы тёмными. Видимо поэтому, человек, в отличие от некоторых насекомых, их не воспринимает.
Инфракрасные (ИК) лучи для жизнедеятельности организма так-же малоинформативны. Они позволили бы человеку различать, например, различные виды белков, но это не столь важно. Ещё одна особенность, ИК лучи приходят на землю от нашего светила — Солнца. Частично они поглощаются атмосферой, при этом спектральный состав дошедших до земли лучей зависит от её состояния и беспрерывно меняется. Изменения погоды привели бы к тому, что цвет предметов беспрерывно менялся, и вряд ли организм сумел бы разобраться в этом хаосе.
Иначе обстоит дело с видимой областью спектра. Это информация о том, спелые плоды или нет, можно ли их съесть или следует воздержаться. То есть, это большая часть необходимой живому организму информации. Света достаточно, атмосфера его почти не поглощает, а его спектральный состав в течение дня почти не меняется. Красный плод остается красным весь день и даже на следующий. Свет — это привязка к источнику жизни, к Солнцу.
Итак, самым информативным участком спектра является видимый. Но так как он очень узок — видимая область занимает участок всего лишь от 380 нанометров (или 0,38 миллионной доли метра) до 780 нанометров, — природа вынуждена была использовать его максимально полно. Для этого она разукрасила его всеми цветами радуги.
Большинство организмов, даже простейших, реагируют на свет. С развитием живых существ развивался и механизм восприятия ими света. В дальнейшем этот механизм преобразовался в полноценный орган позволяющий не только определять наличие и направление света, но и воспринимать очертания окружающего мира. Этим органом стал глаз. С развитием организмов совершенствовался и сам глаз. У более высокоразвитых существ, орган зрения позволяет не только видеть окружающее пространство, но и различать предметы по их способности поглощать/отражать различные участки солнечного спектра (восновном в так называемой видимой области). При этом глаз стал различать цвета предметов.
Изначально рецепторы были простейшими, реагирующими восновном на определённую длину волны. При этом орган зрения позволял различать достаточно широкую гамму оттенков по яркости. У приматов (и человека) мутация вызвала появление колбочек — цветовых рецепторов. Это было вызвано расширением экологической ниши млекопитающих, переходом части видов к дневному образу жизни.
Несмотря на широко распространенное убеждение, что цветовое зрение — редкость у млекопитающих, большинство которых якобы видит только оттенки серого, судя по накапливающимся фактам, многие виды, включая домашних кошек и собак, все же, хотя бы в некоторой степени, различают и цвета. Цветовое зрение, вероятно, наиболее развито у приматов, но известно также у лошади, жирафа, виргинского опоссума, нескольких видов белок и многих других зверей.[1]
Эволюция цветного зрения (см. цветное зрение) связана как с эволюционной адаптацией зрения, как рецепторной системы, так и с влиянием благоприятных и неблагоприятных мутаций.
Доступное нашему зрению многообразие различных цветов определяется наличием опсинов — светочувствительных белков, которые синтезируются в фоточувствительных рецепторах глаза. Опсины принадлежат к семейству ретинолидных белков, которые обнаружены в светочувствительных клетках сетчатки. Классифицируют два типа опсинов — первого и второго типов. Сходство в их структуре и функции указывает на их конвергентное происхождение у бактерии и животных.[2]
В опсинах прокариот различают семь трансмембранных доменов, сходные по структуре с рецепторами, связанными с G-белками у эукариот. Несмотря на определённое сходство их строения, нет никаких подтверждений того, что они имеют эволюционное сходство; по-видимому, эти белки эволюционировали независимо друг от друга.[2]
Некоторые бактерии фиксируют углерод без участия хлорофилла, используя для этого опсины первого типа (протеородопсины, галородопсины и бактериородопсины) Галобактерии используют фоточувствительные родопсины в процессах фототаксиса, в этом процессе участвуют, связанные с плазматическими мембранами, белки системы передачи сигнала, не имеющие отношения к G-белкам.[3]
Два семейства опсинов позвоночных классифицируют по различиям в экспрессии и эволюционной истории. Родопсины, которые активно участвуют в сумеречном зрении, являются высокочувствительными к свету опсинами, и располагаются в палочках сетчатки, но с относительно невысокой плотностью, не обеспечивающей достаточной разрешающий способности глаза. Опсины колбочек расположены в клетках колбочек сетчатки, они участвуют в цветном зрении, и имеют меньшую светочувствительность, чем опсины палочек. Зато их плотность в сетчатке выше, поэтому они обеспечивают высокую разрешающую способность дневного зрения. Опсины колбочек также классифицируют по длине волн максимального поглощения света. Также для классификации часто сравнивают аминокислотные последовательности опсинов разных групп.
Оба подхода позволяют выделить четыре основных группы опсинов.[4] У человека представлен следующий набор зрительных белков:
- Родопсин (Rh1, OPN2, RHO) — экспрессируется в палочках, участвует в ночном зрении
- Четыре типа опсинов колбочек (фотопсины) — экспрессируются в колбочках, участвуют в цветном зрении
- Чувствительные в области длинных волн (Long Wavelength Sensitive, LWS, OPN1LW) опсины — λmax в красном диапазоне
- Чувствительные в области волн средней длины (Middle Wavelength Sensitive, RH2 или MWS) опсины — λmax в зеленой части спектра
- Чувствительные в области коротких волн 2 (SWS2) опсины — λmax в синем диапазоне
- Чувствительные в области коротких волн 1 (SWS1) опсины — λmax в фиолетовой и ультрафиолетовой части диапазона
Для последних трех опсинов у человека известны связанные с ними гены: OPN1MW, OPN1MW2, и OPN1SW.
{kind=link}
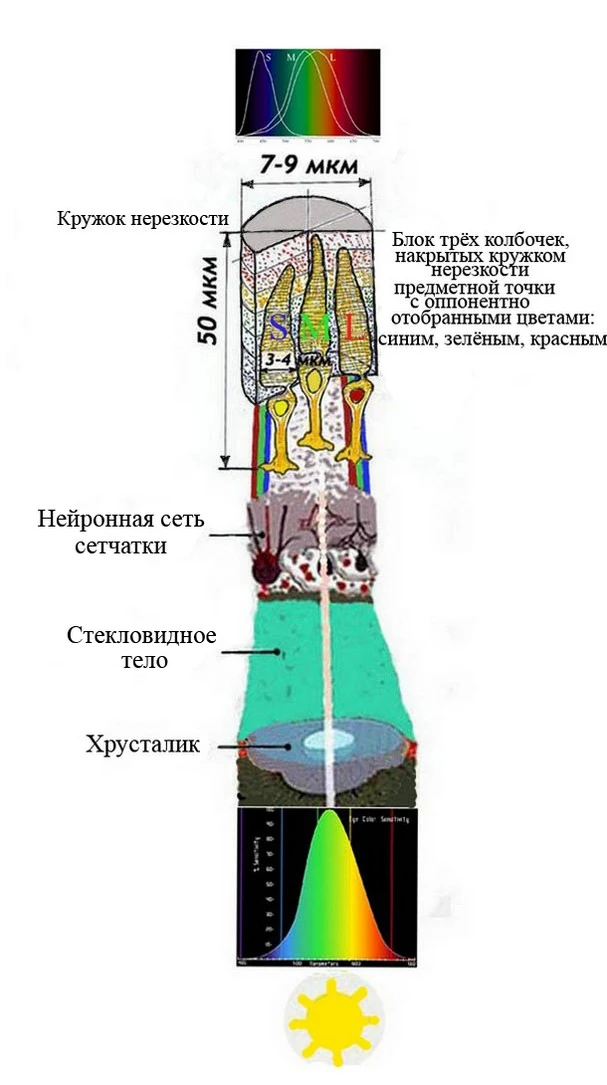
Принципиальная схема цвветного зрения человека, приматов трихроматизма с оппонентным отбором основных цветов предметной точки в условиях ретиномоторной реакции фоторецепторов в блоках колбочек RGB
В настоящее время считается, что наличие определённого числа опсинов в сетчатке животного должно означать и существование такого же числа колбочек (видимо исходя из предположения: одна колбочка — один опсин). С точки зрения рациональности работы колбочек в системе мозаики сетчатки, где колбочки группируются блоками, например, по три или четыре с со своими разновидностями пигмета опсина, воспринимающих каждой колбочкой свои цвета. Рентгеноскопия среза сетчатки птиц, показывает блок из четырёх колбочек фиолетовой, синей, зелёной, красной, который накрывается кружком нерезкости предметной точки.При этом сфокусированный на фокальной поверхности сетчатки размером 7-9 мкм, множество моноточек по нервным стволам от синапса колбочек свои сигналы без изменения угла прохождения, по типу волновода попадают в мембраны колбочек. Здесь оппонентно отбирается более сильный сигнал одного цвета. Например, красная колбочка отбирает красные лучи, зелёная — зелёные и т.д.. Это зависит от вида пигмента. Если принять, что в колбочке присутствуют одновременно все пигметны, то все колбочки не смогут оппонентно отобрать свой наиболее яркий цвет. Они дадут одинаковые усредненные сигналы одного смеешанного цвета. Ведь кружок нерезкости при фокусировании содержит множество спактральных точек, равномерно распределённых в обьёме цилиндра высотой мембраны, в блоке колбочек. (см. рис.). Имелась попытка доказатать в 1964 году[5], что колбочки все одинаковые и содержат набор всех пигментов, откуда нет необходитости их разделять на разные цвета. Однако, параллельно учёными ёщё с 1966 на базе рентгеноскопических исследований доказано, что основой цветного зрения является, например, система трихроматизма трёх колбочек синей, зелёной, красной, содержащих свои пигменты [6].(См. также Лаборатория Р.Е.Марка, Цветное зрение у птиц).
Если в сетчатке глаза отсутствуют или частично вырождены какие либо из фоточувствительных пигментов, то у человека наблюдается цветоаномалия выраженная в неправильной цветопередаче воспринимаемых глазом цветов.
Люди с нормальным цветным зрением имеют в фоторецепторах все три пигмента - разновидностей опсина и в зависимости от вида и структуры пигмента опсина их молекулы максимально чувствительны к длинным длинам волны света (красному цвету), средним длинам волны света (зеленому цвету) или коротким длинам волны света (синему цвету). Откуда, колбочки с различной чувствительностью к спектральным лучам света в зонах (S, M., L — синяя, зелёная, красная) (см. рис. 13) в зависимсти от длины волны и последовательности троп возможности прохождения в мозг сигнала, безусловно, основа цветного восприятия окружающей среды и создания нашего визуального оптического изображения.
В силу морфологических отличий, описанных выше, существуют два основных типа фоторецептора палочек и колбочек, находящиеся в позвоночной сетчатке. Палочки — фоторецепторы, которые содержат зрительный пигмент — родопсин, чувствительный к синему-зеленому цвету с пиковой чувствительностью, равной длине волны света 498 нм. Палочки — очень чувствительные фоторецепторы и используются для видения в условичх сумеречного (скотопического) зрения [7]. В силу ретиномоторной реакции фоторецепторов при достижении освещения 498 нм родопсин обесцвечивается и палочки уходят в зону, где они не принимают дневной свет. Т.е. палочки в цветном зрения не участвуют. Колбочки же содержат пигменты - опсины, и в зависимости от вида и структуры опсина их молекулы максимально чувствительны к длинным длинам волны света (красному цвету), средним длинам волны света (зеленому цвету) или коротким длинам волны света (синему цвету). Колбочки с различной чувствительностью (см. рис. 13) от длины волны и последовательностью троп возможности прохождения в мозг, конечно, основа цветного восприятия окружающей среды и создания нашего визуального оптического изображения. (Здесь особое место занимают колбочки-S)
Сетчатки приматов и человека все еще содержат типы колбочек, которые выглядят по существу аналогично морфологически, но здесь одновременно с последними анатомическими методами исследования замечено, что по крайней мере, наблюдается различие между колбочками с короткой длиной волной и двумя колбочками с более длинными длинами волны. Специализированные гистохимические методы (Марк и Sperling, 1977), исследования с умелым подбором краски (DeMonasterio и др., 1981) или использование определённых антител для визуальных пигментов (Szel и др., 1988), позволили идентификацию различных спектральных (цветовых) типов колбочек теперь наиболее относящихся к разновидностям млекопитающих. В антителах сетчатки примата противоположные визуальные пигменты окрашивают внешние доли L/M-колбочек вместе, в то время как Булочек-S-колбочек окрашивают самостоятельно.
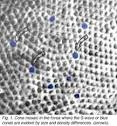
{kind=link}
Рис.1. Мозаика колбочки в ямке, где S-волна или синие колбочки очевидны различиями плотности и размером.[8]
В вышеупомянутом антивизуальном антителе пигмента запятнанная ткань, Булочки-колбочки-S выделяется в виде колбочек, которые не запятнаны, потому что антитело признает только колбочки L и М (оппонентно). Т.е. фотопигмент окрашивает в виде коричневых запятнанных профилей колбочек типов - L-и М, в то время как незапятнанные профили, окруженные синими кругами - это Булочки или колбочки-S (Wikler и Rakic, 1990). Т.о. не может быть речи о причастности родопсина (палочек) в цветном зрении (трихроматизма).
В этой связи аномалии цветного зрения как дальтонизм связаны с отсутствием или нарушением работы фототрансдукции сигнала S,M,L пигментов колбочек того или иного цвета. Это прежде всего пигментов йодопсинов, кон-опсинов, меланопсина (см. опсины), ответственных за красно-зелёное восприятие цветов, а также сине-жёлтых цветов.
Известны три частных случая цветоаномалии дальтонизма.
1.Отсутствует пигмент (сенсибилизатор), реагирующий на длинноволновую (жёлто-красную) область спектра-L, - разновидность эритролаб. Этот дефект называют дальтонизмом 1-го рода — протанопия.
2.Отсутствует пигмент реагирующий в основном на жёлто-зелёную область спектра-M — хлоролаб. Такое цветовосприятие свойственно при дальтонизме 2-го рода - дейтеранопия.
3.Отсутствует пигмент, реагирующий в основном на фиолето-синюю область спектра-S — разновидность йодопсина (Булочки-колбочки-S) - так называемая "куриная слепота". При этом помимо искажения цветопередачи у человека отсутствует сумеречное зрение. В данном случае также нарушена функция работы фоторецепторов палочек, содержащих фотопигмент родопсин [9] реагирующий на фиолетово-синий спект света. Этот случай и есть дальтонизм 3-го рода - тританопия (см.Цветное зрение у птиц, Лаборатория Р.Е.Марка, Участник:Миг/Тропы булочек (S-cone)).
Редко встречается парная, или даже полная цветовая слепота.
Последние данные исследований: см.в Цветное зрение у птиц, в отчёте Лаборатория Р.Е.Марка, доказали , что колбочки в блоках имеют разные цвета и реагируют на один свой цвет.
Несмотря на это, многие продолжают считать, что большинство млекопитающих обладает двуцветным видением (по количеству обнаруженных в сетчатке опсинов), различающим короткие и длинные диапазоны волн называя их дихроматами. При этом утверждается, что приматы уникальны, поскольку они «трихроматы» — то есть обладают «трёхкомпонентным» зрением, т.е обладают тремя опсинами — чувствительными к разным диапазонам видимого света[10]. en:Platyrrhine. Кроме того, существует и гипотеза, что часть людей, особенно среди женщин могут быть — «тетрахроматы». Чаще всего эти противоречивые догадки связаны с упрощённой интерпритацией процесса зрения и так и невыясненного на сегодя (несмотря на обилие предположений, гипотез и теорий) истинного механизма цветного зрения.
См. также[]
Примечания[]
- ↑ http://www.krugosvet.ru/enc/nauka_i_tehnika/biologiya/MLEKOPITAYUSHCHIE.html#1000247-L-111
- ↑ 2,0 2,1 Fernald, Russell D. (September 2006), "Casting a Genetic Light on the Evolution of Eyes", Science 313 (5795): 1914–1918, doi:, PMID 17008522
- ↑ Römpler H, Stäubert C, Thor D, Schulz A, Hofreiter M, Schöneberg T (February 2007), "G protein-coupled time travel: evolutionary aspects of GPCR research", Molecular Interventions 7 (1): 17-25, doi:, PMID 17339603
- ↑ Terakita A (2005). "The Opsins". Genome Biology 213 (6(3)): 213. doi:10.1186/gb-2005-6-3-213.
- ↑ Marks W. B., W. H. Dobelle, E. F. McNichol. 1964. Science, 143 : 1181.
- ↑ http://www.library.biophys.msu.ru/PDF/3353.pdf М. А. Островский ФОТОБИОЛОГИЧЕСКИЙ ПАРАДОКС ЗРЕНИЯ
- ↑ http://www.library.biophys.msu.ru/PDF/3353.pdf М. А. Островский ФОТОБИОЛОГИЧЕСКИЙ ПАРАДОКС ЗРЕНИЯ
- ↑ http://webvision.med.utah.edu/S-cone.html
- ↑ http://www.library.biophys.msu.ru/PDF/3353.pdf М. А. Островский ФОТОБИОЛОГИЧЕСКИЙ ПАРАДОКС ЗРЕНИЯ
- ↑ Bowmaker, J. K. 1998. Evolution of colour vision in vertebrates. Eye. 12: 541—547.